Влияние присутствия токсичных добавок в составе полимерного материала на процессы его биодеструкции в морской воде
Проведена экспозиция в течение 60 сут образцов отвержденной полиэфирной смолы с добавлением токсичных оксидов металлов и целлюлозы (контрольный образец) в морской воде Геленджикской бухты Черного моря.Показано, что эукариотные диатомовые водоросли, а также прокариотные сине-зеленые водоросли (цианобактерии) были наиболее многочисленными членами сообществ, заселяющих поверхности образцов. Методом секвенирования V4 региона гена 16S рРНК прокариот и биоинформатического анализа этих результатов показано, что в обрастаниях на образцах с оксидами хрома, свинца, цинка и титана возрастала доля бактерий, устойчивых к этим металлам, тогда как бактерии, потенциально способные деградировать полиэфирные смолы, составляли минорную часть сообществ.
Введение
В последние десятилетия разработаны материалы, максимально стойкие к воздействию агрессивных сред, что предполагает насущную необходимость создания способов их утилизации с минимальным ущербом для окружающей среды [1].
Устойчивость полимерных материалов к биодеструкции зависит от различных факторов: шероховатости поверхности, химической структуры и физического состояния полимеров, а также физико-химических условий среды, в которой они эксплуатируются [2–4]. Полимеры с гидролизуемыми химическими связями в основной цепи, такие как полиэтилентерефталат, более подвержены биодеградации [5], чем такие, как полистирол, в структуре которого отсутствуют гидролизуемые функциональные группы [6].
В морской воде на поверхности материалов неизбежно формируются обрастания (микробные биопленки). Биопленки усиливают микробную активность, защищают микроорганизмы от ультрафиолета, радиации, высокой солености, тяжелых металлов и антибиотиков [7, 8]. Однако пока отсутствует понимание, чем в большей степени определяется состав формирующихся биопленок – физико-химическими факторами и микрофлорой окружающей водной среды или свойствами экспонируемых материалов. Имеющиеся в научно-технической литературе данные, полученные в результате изучения биопленок, развивающихся на различных материалах в водных средах, пока не позволяют однозначно ответить на этот вопрос. В частности, установлено, что микробные скопления на пластике и стекле формируются не в зависимости от поверхностных свойств материала, а под влиянием места экспозиции и сезонности [9, 10].
В предыдущих работах авторами показано заметное влияние характера водной среды обитания на состав микробных сообществ обрастаний [11].
Полиэфирные смолы широко производят в виде полимерных композиционных материалов для изготовления корпусов легких судов, буев и других изделий. Такие изделия из полиэфирной смолы длительно контактируют с морской водой и морской микрофлорой, поэтому обеспечение их длительной работоспособности при воздействии влаги и микроорганизмов является актуальной задачей.
Цель данной работы – определение состава бактериальных сообществ обрастаний на образцах отвержденной полиэфирной смолы, содержащей наполнители в виде оксидов металлов различной токсичности, в морской воде Геленджикской бухты Черного моря.
Материалы и методы
Образцы изготовлены из прозрачной полиэфирной смолы для литья марки Norsodyne O 12335 AL, отвержденной отвердителем Бутанокс М50. Для смазки формы использовали восковую разделительную спрей-смазку марки Вс-М. Добавляли наполнители в количестве 10 % (по массе) (рис. 1, а). Образцы размещали на литорали побережья Черного моря в районе г. Геленджика (рис. 1, б). На основании проведенных экспериментов установлено, что заметное обрастание образцов происходило только после 15 сут экспозиции, а полное обрастание – после 60 сут. Поэтому образцы для анализа снимали через 60 сут экспозиции в августе 2020 г. (рис. 1, в). Ионный состав морской воды в месте экспонирования приведен в табл. 1.

Рис. 1. Подготовка и экспозиция образцов: а – образцы полиэфирной смолы с добавлением соединений металлов различной токсичности в форме для отливки; б – экспозиция образцов в морской воде; в – внешний вид образцов после окончания экспозиции в Черном море
Таблица 1
Ионный состав морской воды в точке экспонирования
(средние значения и диапазоны полученных значений за период экспозиции образцов)
Катионы, мг/л | Анионы, мг/л | |||
K+ | Na+ | Mg2+ | Cl– | |
65–170 130 | 2608–5058 4052 | 282–622 500 | 5918–10890 9396 | 856–1487 1277 |
* В числителе – минимальное и максимальное значения, в знаменателе – среднее. | ||||
Методы анализа состава прокариот в обрастаниях на полимерных материалах
Состав микробных сообществ определяли путем синтеза ампликонов гена 16S рРНК прокариотических организмов и последующей биоинформатической обработки полученных данных. Ампликоны гена 16S рРНК получали путем полимеразной цепной реакции с использованием препаратов тотальной дезоксирибонуклеиновой кислоты (ДНК), выделенной из микробных обрастаний на анализируемых образцах.
Выделение ДНК из образцов (фильтров) осуществляли с использованием коммерческого набора для выделения ДНК FastDNA Spin Kit (фирма MP Bio, США) в соответствии с инструкцией производителя. Cоотношение OD 260/280 в полученном препарате ДНК – не менее 1,8. Измерение массовой концентрации ДНК осуществляли флуориметрическим методом. Соотношение OD 260/280 (отношение значения оптической плотности при длине волны пропускаемого света 260 нм к значению оптической плотности при длине волны пропускаемого света 280 нм) измеряли с использованием спектрофотометра. Далее полученный препарат ДНК использовали для амплификации V4 участка гена 16S рРНК путем полимеразной цепной реакции. Получаемые при этом ампликоны за счет наличия на 5ʹ-концах праймеров технических последовательностей могут быть после очистки секвенированы на платформе Illumina MiSeq. Технические последовательности включали в себя адапторы Illumina, сайты посадки секвенирующих праймеров, баркоды и спейсеры гетерогенности. Очистку ампликонов осуществляли путем препаративного агарозного гель-электрофореза и последующего использования стандартных наборов реактивов для очистки ампликонов из агарозного геля и реакционных смесей со стандартными спин-колонками в соответствии с рекомендациями производителей наборов. Очищенные и готовые для секвенирования ампликоны подготавливали для секвенирования на платформе Illumina MiSeq в соответствии с рекомендациями производителя.
Приготовление библиотек гена 16S рРНК
для высокопроизводительного секвенирования на системе Illumina MiSeq
Библиотеки гена 16S рРНК для высокопроизводительного секвенирования на системе Illumina MiSeq приготовлены по схеме, описанной в статье [12]. Для получения ампликонов V4 использовали следующую систему праймеров: прямой праймер 5′ Illumina Linker Sequence (Index 1, Heterogeneity Spacer) [13] и 515F праймерной последовательности [14]; обратный праймер 3′ Illumina Linker Sequence (Index 2, Heterogeneity Spacer) и Pro-mod-805R праймерной последовательности [15] соответственно.
Высокопроизводительное секвенирование библиотек гена 16S рРНК
и первичная обработка полученных данных
Высокопроизводительное секвенирование полученных на предыдущей стадии библиотек выполняли с использованием набора реагентов MiSeq Reagent Micro Kit v2 (300-cycles) MS-103-1002 (компания Illumina, США) на секвенаторе MiSeq (компания Illumina, США) в соответствии с инструкцией производителя. Предварительная биоинформатическая обработка заключалась в объединении прямых и обратных прочтений, фильтрации последовательностей с низкими показателями достоверности прочтения отдельных нуклеотидов, фильтрации химерных последовательностей, распределении прочтений по образцам на основании последовательностей баркодов и удалении технических последовательностей (в том числе и праймерных последовательностей на V4 фрагмент гена 16S рРНК). Затем полученные последовательности распределяли по операционным таксономическим единицам (ОТЕ) с уровнем сходства последовательностей >97 %. Полученные ОТЕ биоинформатическим путем идентифицировали на основании базы данных Silva. Относительное количество прочтений, входящих в ту или иную ОТЕ, соответствует относительной представленности данного таксона микроорганизмов в исследуемом микробном сообществе [16–18].
Библиотеки фрагментов гена 16S рРНК из микробных обрастаний на образцах смолы с наполнителями, полученные в результате высокопроизводительного секвенирования, депонированы в NCBI SRA биопроект PRJNA856151.
Результаты и обсуждение
Статистический анализ разнообразия микроорганизмов
в обрастаниях на образцах полиэфирной смолы с наполнителями
В результате высокопроизводительного секвенирования V4 региона гена 16S рРНК прокариот получено шесть библиотек, содержащих 171049 прочтений (ридов), из которых сформировано 12180 ОТЕ (филотипа) с уровнем сходства последовательностей ≥97 % (табл. 2).
Уровень покрытия для всех библиотек был относительно невысоким и составлял от 80 до 87 %, что свидетельствовало о том, что максимальная репрезентативность библиотек не была достигнута. Этому заключению соответствовали и высокие значения индекса Chao 1. Однако такой уровень покрытия позволяет достаточно достоверно количественно выявлять все доминирующие таксоны микроорганизмов.
Индекс разнообразия Шеннона–Уивера оказался заметно выше для контрольного образца с целлюлозой и образцов с титаном, чем для образцов с цинком, хромом и свинцом. Индекс Бергера–Паркера, наоборот, был более низким для образцов с целлюлозой и титаном по сравнению с образцами с цинком, хромом и свинцом, что свидетельствовало о более высоком разнообразии микроорганизмов, т. е. добавление ZnO, PbO2 и Cr2O3 к полимерам способствовало снижению разнообразия микроорганизмов в биообрастаниях на этих полимерах.
Таблица 2
Индексы разнообразия генов 16S рРНК бактерий в обрастаниях
на образцах полиэфирной смолы с наполнителями
Параметры | Библиотеки фрагментов гена 16S рРНК | |||||
VIAM1L20 | VIAM2L20 | VIAM3L20 | VIAM4L20 | VIAM6L20 | VIAM7L20 | |
Количество сиквенсов (ридов) | 24484 | 27878 | 30993 | 26329 | 31486 | 29879 |
Количество ОТЕ* | 3241 | 3166 | 3901 | 3918 | 4229 | 3439 |
Покрытие по Good, % | 83 | 87 | 83 | 80 | 83 | 87 |
Индекс Chao 1 | 7545 | 7918 | 9425 | 10274 | 10265 | 8459 |
Индекс разнообразия Шеннона–Уивера (H) | 6,29 | 5,26 | 5,78 | 6,43 | 6,51 | 5,83 |
Индекс доминирования Бергера–Паркера (D) | 0,03 | 0,20 | 0,25 | 0,04 | 0,04 | 0,13 |
* Общее количество ОТЕ в библиотеках меньше суммы ОТЕ в каждой библиотеке, потому что значительное количество ОТЕ являются общими для части или всех библиотек. | ||||||
Группирование библиотек на уровне OTE методом главных компонент (рис. 2) показало обособленное положение контрольного образца с целлюлозой, а также отдаленное положение двух образцов с титаном от группы образцов смол с цинком, хромом и свинцом.
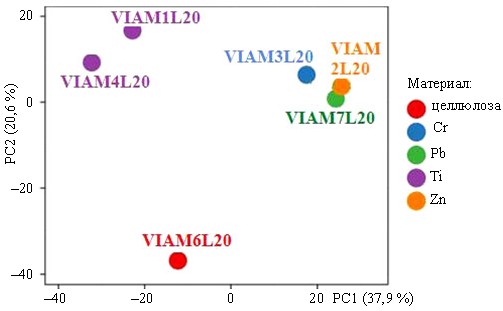
Рис. 2. График группирования таксономических единиц (ОТЕ) в библиотеках фрагментов генов бактерий и грибов по материалу наполнения образцов – титан (Ti), цинк (Zn), хром (Cr), свинец (Pb) и древесные опилки (целлюлоза), полученный методом главных компонент РС1 и РС2 (Principal Components Analysis – PCA) с использованием параметра разложения по сингулярным значениям в программе ClustVis
Таксономический состав микроорганизмов в обрастаниях
на образцах полиэфирной смолы с наполнителями
Состав высших таксонов бактерий сходен для всех анализируемых библиотек, при этом представленными в библиотеках и доминирующими в большинстве из них были последовательности бактерий филумов/классов Alphaproteobacteria,Gammaproteobacteria, Bacteroidota, хлоропластов эукариотных диатомовых водорослей (Bacillariophyta), Actinomycetota и Cyanobacteria (прокариотных сине-зеленых водорослей) (рис. 3). Однако наблюдались заметные количественные изменения в составе доминирующих высших таксонов, связанные с природой материала наполнителя.
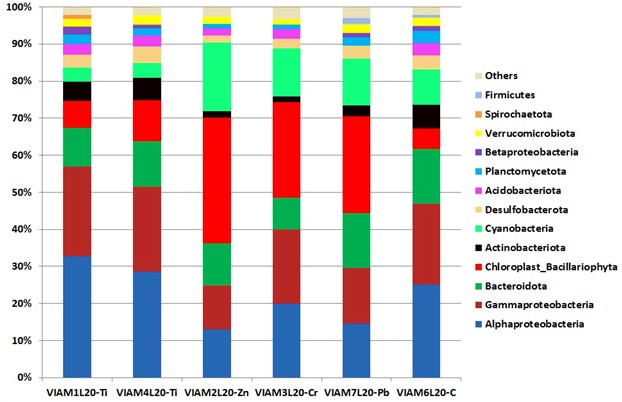
Рис. 3. Относительная доля фрагментов гена 16S рРНК бактерий на уровне филумов/классов в библиотеках из обрастаний на образцах полиэфирной смолы с различными токсичными наполнителями и целлюлозой (С), экспонированных в морской воде. Представлены таксоны, составляющие не менее 1 % в каждой библиотеке
Для библиотек контрольного образца с древесными опилками и с оксидом титана характерно преобладание бактерий класса Alphaproteobacteria (24,2–32,7 %), среди которых много представителей типичной морской микрофлоры. Различием в этой группе было более низкое содержание в Ti-библиотеках цианобактерий (3,8–4,0 %) по сравнению с библиотекой контрольного образца (9,6 %). В то же время в библиотеках обрастаний на образцах смолы с оксидами цинка, хрома и свинца доля Alphaproteobacteria была заметно меньше (13,0–20,0 %), при этом преобладали хлоропласты Bacillariophyta (25,7–34,0 %) и цианобактерии (12,6–18,4 %). Таким образом, основным компонентом обрастаний на поверхностях образцов с цинком, хромом и свинцом были фотоавтотрофные эу- и прокариотные микроорганизмы, выделяющие кислород.
В результате последующей классификации ОТЕ в общем составе шести библиотек фрагментов гена 16S рРНК выявлены представители 512 родов бактерий, включая неидентифицированные цианобактерии, а также хлоропластыэукариотных диатомовых водорослей Bacillariophyta. На рис. 3 приведены преобладающие таксоны родового уровня, последовательности которых составляли не менее 1 % прочтений в каждой библиотеке. В библиотеке контрольного образца с целлюлозой представлено 17 доминирующих родов, среди которых преобладали неидентифицированные цианобактерии (8,2 %) и бактерии рода Fulvivirga (филум Bacteroidota) (3,0 %); хлоропласты Bacillariophyta составляли 5,4 %. Бактерии остальных родов составляли от 1 до 2,5 % каждый от общего состава библиотеки. Похожий уровень разнообразия родов бактерий представлен в обеих библиотеках с титановым наполнителем (по 24 рода), в которых преобладали хлоропласты Bacillariophyta (7,3–11,1 %), а филотипы бактерий составляли от 1 до 2,9 % от общего состава каждой из библиотек. Набор доминирующих родов был похожим для обеих Ti-библиотек и их контроля, но при этом обогащен представителями типичной морской микрофлоры, что свидетельствовало о незначительном влиянии на него (набор) материала образцов и наполнителей в виде целлюлозы и оксида титана.
В то же время состав доминирующих родов в библиотеках с оксидами хрома, свинца и цинка гораздо менее разнообразен, чем вышеприведенные, которые содержали всего 6 родов для образца смолы с оксидом цинка и по 8 родов для образцов с оксидами хрома и свинца. В составе этих библиотек выявлено подавляющее преобладание хлоропластов эукариот Bacillariophyta (25,5–33,7 %) и неидентифицированных цианобактерий (11,5–16,2 %), а остальные рода составляли от 1,0 до 4,0 %. Редуцированный состав микробных сообществ на образцах полиэфирных смол с оксидами цинка, хрома и свинца, по-видимому, связан с токсичностью этих наполнителей для микрофлоры обрастаний.
Сопоставление токсичности оксидов металлов
в исследуемых образцах полимерной смолы
При создании авиационной и морской техники используется большое количество материалов и металлических сплавов: сталь, хромоникелевые, алюминиевые, титановые и цинксодержащие сплавы. Все эти металлы имеют различную токсичность по отношению к живым организмам. В табл. 3 приведены сравнительные данные о токсичности наполнителей, использованных в данном исследовании, согласно Гигиеническими нормативами СанПиН 1.2.3685–21 «Предельно допустимые концентрации (ПДК) вредных веществ в воздухе рабочей зоны». Как видно из данных табл. 3, наиболее опасными являются оксиды хрома и свинца, менее опасным – оксид цинка, а малоопасным – оксид титана. Однако эти нормативы получены на опытах с мелкими животными и рыбами, и для бактерий могут быть использованы только как справочный материал.
Таблица 3
Токсичность наполнителей
Наполнитель | Величина ПДК, мг/м* | Класс опасности** | Особенности действия на организм |
Оксид цинка | 1,5/0,5 | 2 | – |
Свинец и его неорганические соединения/по свинцу | –/0,05 | 1 | – |
Триоксид хрома (VI) | 0,03/0,01 | 1 | Промышленные канцерогены |
Диоксид титана | –/10 | 4 | Аэрозоли преимущественно фиброгенного действия |
* Одно значение гигиенического норматива, приведенное в графе, соответствует максимальной разовой предельно допустимой концентрации вещества в воздухе рабочей зоны (ПДК м. р.); если приведено два значения гигиенических нормативов, то это означает, что в числителе стоит значение максимальной разовой (м. р.) предельно допустимой концентрации (ПДК м. р.), а в знаменателе – среднесменной предельно допустимой концентрации (ПДК с. с). ** Химические вещества по величине гигиенических нормативов разделены на четыре класса опасности: | |||
В настоящее время ведутся активные исследования в области устойчивости микроорганизмов к тяжелым металлам. Известно, что значительный бактерицидный эффект может дать взаимодействие оксида титана с ультрафиолетовым облучением, в результате которого образуются свободные радикалы, разрушающие клетки патогенных микроорганизмов [19, 20]. Однако в составе образца полиэфирной смолы, глубоко погруженного в морскую воду, такой эффект вряд ли возможен. С этим согласуется обнаруженное авторами на поверхности образца с титановым наполнителем таксономически разнообразное микробное обрастание (рис. 4, а, б), сравнимое с контрольным образцом с древесными опилками (рис. 4, е). Это свидетельствует о низкой токсичности оксида титана в образце полиэфирной смолы и об отсутствии его бактерицидного эффекта в данных условиях.
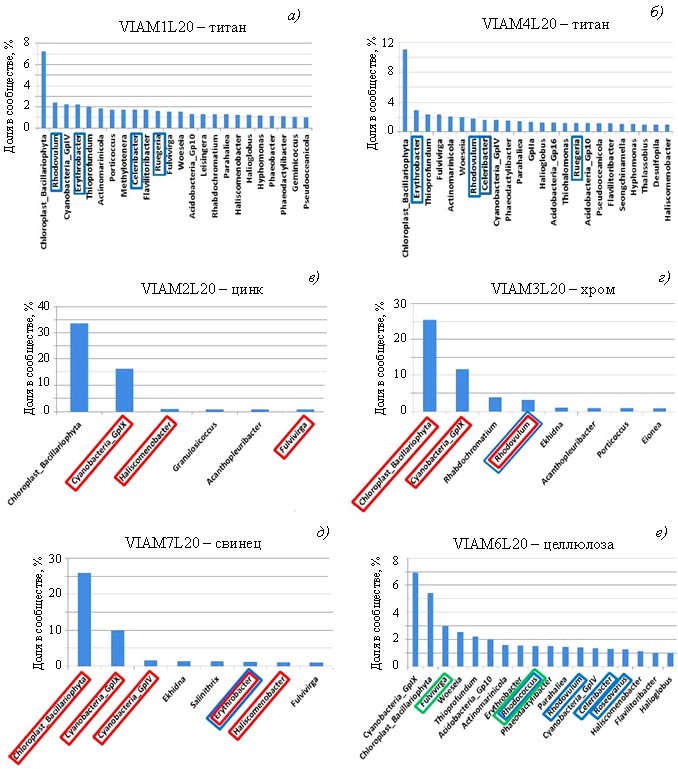
Рис. 4. Относительная доля фрагментов гена 16S рРНК бактерий на уровне родов и классов/порядков (для некультивируемых бактерий) в библиотеках из обрастаний на образцах полиэфирной смолы с различными токсичными наполнителями, экспонированных в морской воде. Представлены таксоны, составляющие не менее 1 % в каждой библиотеке. Красной рамкой выделены таксоны, в геноме которых предполагается наличие генов устойчивости к тяжелым металлам (цинк, хром, свинец). Синей рамкой выделены таксоны, в составе которых предполагается наличие бактерий-деструкторов полимеров (согласно базе данных KEGG). Зеленой рамкой выделены таксоны, в составе которых предполагаются бактерии-деструкторы целлюлозы
Таксономическое разнообразие прокариот на обрастаниях смол с оксидами цинка, хрома и свинца значительно ниже, чем у контрольного образца с целлюлозой, что, по-видимому, связано с отбором резистентной к тяжелым металлам морской микрофлоры. Для многих бактерий обычным способом обеспечения устойчивости к тяжелым металлам является снижение их концентрации с помощью ферментных систем эффузионного оттока ионов металлов из клеток. Эти ферментные системы детерминируются специальными генами, обычно собранными в кластеры и расположенными на внутриклеточных плазмидах или в хромосомах [21].
С помощью базы данных KEGG выполнен сравнительный анализ состава микроорганизмов, доминирующих на образцах с наполнителями из тяжелых металлов, и определены бактерии, в геномах которых предположительно присутствуют гены устойчивости к исследуемым тяжелым металлам. Цинк является мощным ингибитором системы дыхательного транспорта электронов бактерий и митохондрий эукариот. Гены устойчивости к цинку объединены в czc-оперон, который также обеспечивает устойчивость к кадмию и кобальту [22]. В результате сравнительного биоинформатического анализа гены czc-системы обнаружены в геномах различных цианобактерий, а также у бактерий родов Haliscomenobacter и Fulvivirga, которые наиболее представлены в микробном сообществе на образце с оксидом цинка (рис. 4, в). Гены устойчивости к оксиду хрома, который производит окислительные повреждения белков клетки, объединены в chr-оперон [23]. Гены системы устойчивости к хрому найдены в геномах некоторых диатомовых водорослей (Bacillariophyta), различных цианобактерий, а также фототрофных бактерий рода Rhodovulum, доминирующих в сообществе на образце с оксидом хрома (рис. 4, г). Токсичность свинца выражается в повреждениях многих внутриклеточных процессов. Удаление свинца из клеток бактерий осуществляется ферментным комплексом, детерминируемым генами pbr-системы [24]. Такие ферментные системы предположительно присутствовали в геномах некоторых диатомовых водорослей (Bacillariophyta), различных цианобактерий, а также бактерий родов Erythrobacter и Haliscomenobacter (рис. 4, д).
Таким образом, можно предположить, что уменьшение таксономического разнообразия в обрастаниях на образцах полиэфирной смолы с наполнителями тяжелых металлов связно с токсичностью последних. При этом в составе доминирующих в обрастаниях таксонов предположительно присутствуют микроорганизмы, обладающие ферментными системами устойчивости к тяжелым металлам.
Потенциальные бактерии-деструкторы полимеров
в обрастаниях на исследуемых образцах полиэфирной смолы
Используемая в данном исследовании полиэфирная смола относится к ненасыщенным смолам, являющимся результатом конденсации этиленгликоля с терефталевой кислотой, с растворением в стироле [25].
Программа iVikodak позволяет на основании таксономического состава (на уровне рода) исследуемых бактериальных сообществ предсказывать их функциональные характеристики, используя геномные базы данных. С использованием модуля программы Global Mapper на рис. 5 представлен результат сравнительного анализа вклада исследуемых бактериальных сообществ в осуществление основных путей метаболизма, а также путей деградации химических соединений-ксенобиотиков на основании базы данных KEGG. Согласно полученным результатам, наибольшим потенциальным функциональным разнообразием для всех путей метаболизма отличались сообщества Ti-образцов и контрольного образца с целлюлозой, что согласуется с большим таксономическим разнообразием этих библиотек.
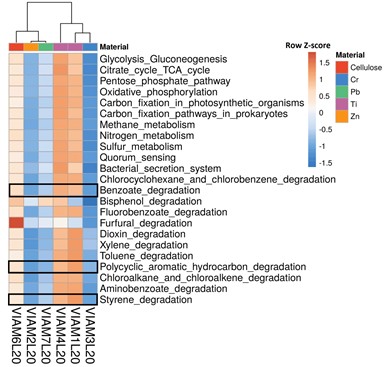
Рис. 5. Результат сравнительного анализа вклада исследуемых бактериальных сообществ
в осуществление основных путей метаболизма, а также путей деградации химических соединений-ксенобиотиков на основании базы данных KEGG
С помощью следующего модуля программы Local Mapper на основании родового состава сообществ, сравнительного анализа геномов из базы данных KEGG для выявленных родов, а также представленности в этих геномах генов (ферментов) различных путей метаболизма возможно получение предположительной оценки вклада различных родов бактерий в эти процессы. При этом согласно результатам программы iVikodak, состав наиболее вероятных бактерий-деструкторов не обязательно совпадает с составом количественно доминирующих в сообществах родов.
Ферменты метаболизма этиленгликоля представлены в составе KEGG-пути «Метаболизм глиоксилата и дикарбоксилата», среди которых ключевым является лактальдегидредуктаза. Основной вклад в деградацию этиленгликоля предположительно осуществляли бактерии родов Rhodovulum, Roseovariusи Rhodococcus(табл. 4).
В составе пути метаболизма «Деградация полициклических ароматических углеводородов» представлены ферменты деградации фталата с последующим включением продукта его деградации через путь «Деградация бензоата» в клеточный метаболизм. Основной вклад в осуществление этих путей вносили альфапротеобактерии родов Rhodovulum, Roseovarius,Ruegeria,Celeribacter,Erythrobacter,а также актинобактерии родаRhodococcus. Точно такой же состав предположительных деструкторов стирола выявлен программой при анализе ферментов, представленных в составе пути «Деградация стирола». Основной вклад в деградацию целлюлозы в составе пути «Метаболизм крахмала и сахарозы» в контрольном образце вносили бактерии родов Fulvivirga и Rhodococcus(рис. 4, е).
Сравнительный анализ присутствия потенциальных бактерий-деструкторов компонентов полиэфирной смолы в составе микробных сообществ на различных исследуемых образцах показал, что они представлены среди количественно доминирующих в сообществах родов преимущественно на образцах с целлюлозой и оксидом титана (рис. 4, а, б, е). Исключение составляли только бактерии рода Rhodovulumна образце смолы с хромом (рис. 4, г) и рода Erythrobacter– на образце смолы со свинцом (рис. 4, д). Известно, что бактерии этих родов обладают генами устойчивости к соответствующим металлам. Таким образом, можно предположить, что добавки тяжелых металлов к образцам полиэфирной смолы уменьшают долю потенциальных бактерий-деструкторов в обрастаниях и повышают устойчивость полимерного материала к биодеструкции, особенно с добавками ZnO.
Таблица 4
Рода бактерий*, потенциальных деструкторов ксенобиотиков
и компонентов полимерных материалов, согласно программе iVikodak
Библиотека (наполнитель) | Пути метаболизма по базе данных KEGG и разлагаемые соединения, предсказанные программой iVikodak (предположительный вклад в процесс биодеструкции) | |||
Метаболизм глиоксилата и дикарбоксилата (этиленгликоль) | Деградация полициклических ароматических углеводородов (фталат) | Деградация бензоата (бензоат, катехол, бензол) | Деградация стирола (стирол) | |
VIAM1L20: VIAM4L20 (Ti) | Rhodovulum (6,3 %) | Celeribacter (12,0 %); Rhodovulum (10,5 %); Ruegeria (5,1 %) | Rhodovulum (7,3 %); Erythrobacter (6,1 %) | Erythrobacter (6,0 %); Celeribacter (5,3 %); Ruegeria (5,3 %); Rhodovulum (5,1 %) |
VIAM2L20 (Zn) | Roseovarius (5,6 %); Rhodovulum (5,4 %) | Celeribacter (9,6 %); Rhodovulum (9,1 %); Roseovarius (7,2 %); Rhodovulum (5,1 %) | Rhodovulum (6,2 %) | Roseovarius (5,1 %) |
VIAM3L20 (Cr) | Rhodovulum (16,7 %) | Rhodovulum (27,1 %); Celeribacter (6,8 %); Ruegeria (5,2 %) | Rhodovulum (19,5 %) | Rhodovulum (14,0 %); Ruegeria (5,7 %) |
VIAM7L20 (Pb) | Rhodococcus (6,5 %) | Rhodococcus (12,2 %); Celeribacter (9,0 %); Rhodovulum (7,4 %); Roseovarius (5,1 %) | Rhodococcus (12,8 %) | Rhodococcus (8,8 %) |
VIAM6L20 (целлюлоза) | Roseovarius (5,1 %); Rhodococcus (5,5 %) | Rhodococcus (12,9 %); Celeribacter (10,2 %); Rhodovulum (7,7 %); Roseovarius (6,1 %) | Rhodococcus (14,0 %); Rhodovulum (5,2 %) | Rhodococcus (9,9 %) |
* В скобках указан предсказанный программой iVikodak вклад бактерий данного рода в осуществление соответствующего пути метаболизма, значащими признаются значения >5 %. | ||||
Заключения
Проведена экспозиция в течение 60 сут образцов отвержденной полиэфирной смолы с добавлением оксидов металлов и целлюлозы в морской воде Геленджикской бухты Черного моря. С использованием ДНК, выделенной из обрастаний на образцах смолы после экспозиции, выполнено секвенирование V4 фрагмента гена 16S рРНК прокариотных организмов и проведена последующая биоинформатическая обработка полученных данных. Показано, что на поверхности образцов наиболее многочисленными были эукариотные диатомовые водоросли (Bacteriophyta) и цианобактерии – прокариотные сине-зеленые водоросли, которые являются фотоавтотрофами и не оказывают воздействия на полиэфирный материал. Наибольшее разнообразие бактерий обнаружено на образцах с добавлением целлюлозы и титана, обладающих минимальной токсичностью. По мере увеличения токсичности металлов снижается разнообразие микроорганизмов, кроме диатомовых водорослей и цианобактерий, однако в обрастаниях возрастает доля бактерий, устойчивых к присутствию металлов в среде, т. е. состав бактерий в обрастаниях предположительно определяется составом микроорганизмов, присутствующих в воде, а не природой материала.
Проведенные исследования показали, что добавление токсичных наполнителей (особенно ZnO) в виде оксидов тяжелых металлов в полиэфирные смолы снижает вероятность их биодеструкции. Использование методов и базы данных по молекулярной экологии позволяет провести более информативную оценку присутствия и активности биодеструкторов в микробных обрастаниях на поверхности полимерных материалов.
Исследование выполнено при поддержке Министерства науки и высшего образования РФ с использованием оборудования ЦКП «Климатические испытания» НИЦ «Курчатовский институт» – ВИАМ.
- Каблов Е.Н., Лаптев А.Б., Прокопенко А.Н., Гуляев А.И. Релаксация полимерных композиционных материалов под длительным действием статической нагрузки и климата (обзор). Часть 1. Связующие // Авиационные материалы и технологии. 2021. № 4 (65). Ст. 08. URL: http://www.journal.viam.ru (дата обращения: 10.05.2022). DOI: 10.18577/2713-0193-2021-0-4-70-80.
- Каблов Е.Н. Материалы нового поколения и цифровые технологии их переработки // Вестник Российской академии наук. 2020. Т. 90. № 4. С. 331–334.
- Каблов Е.Н., Старцев В.О. Системный анализ влияния климата на механические свойства полимерных композиционных материалов по данным отечественных и зарубежных источников (обзор) // Авиационные материалы и технологии. 2018. № 2 (51). С. 47–58. DOI: 10.18577/2071-9140-2018-0-2-47-58.
- Коган А.М., Николаев Е.В., Голубев А.В., Лаптев А.Б., Мовенко Д.А. Этапы биообрастания и коррозии стали в черноморской воде // Труды ВИАМ. 2019. № 6 (78). Ст. 09. URL: http://www.viam-works.ru (дата обращения: 08.06.2022). DOI: 10.18577/2307-6046-2019-0-6-84-94.
- Yoshida S., Hiraga K., Takehana T. et al. A bacterium that degrades and assimilates poly(ethylene terephthalate) // Science. 2016. Vol. 353. P. 759–759. DOI: 10.1126/science.aad6359.
- Krueger M.C., Seiwert B., Prager A. et al. Degradation of polystyrene and selected analogues by biological Fenton chemistry approaches: Opportunities and limitations // Chemosphere. 2017. Vol. 173. P. 520–528. DOI: 10.1016/j.chemosphere.2017.01.089.
- Лаптев А.Б., Николаев Е.В., Куршев Е.В., Горяшник Ю.С. Особенности биодеструкции термопластов на основе полиэфиров в различных климатических зонах // Труды ВИАМ. 2019. № 7 (79). Ст. 10. URL: http://www.viam-works.ru (дата обращения: 08.06.2022). DOI: 10.18577/2307-6046-2019-0-7-84-91.
- De Carvalho C.C.C.R. Marine biofilms: A successful microbial strategy with economic implications // Frontiers of Marine Science. 2018. Vol. 5. Art. 126. DOI: 10.3389/fmars.2018.00126.
- Oberbeckmann S., Kreikemeyer B., Labrenz M. Environmental factors support the formation of specific bacterial assemblages on microplastics // Frontiers in Microbiology. 2018. Vol. 8. Art. 2709. DOI: 10.3389/fmicb.2017.02709.
- Dussud C., Meistertzheim A.L., Conan P. et al. Evidence of niche partitioning among bacteria living on plastics, organic particles and surrounding seawaters // Environment Pollution. 2018. Vol. 236. P. 807–816. DOI: 10.1016/j.envpol.2017.12.027.
- Турова Т.П., Соколова Д.Ш., Назина Т.Н., Груздев Д.С., Лаптев А.Б. Филогенетическое разнообразие микробных сообществ с поверхности полиэтилентерефталатных материалов при экспозиции в водных средах // Микробиология. 2020. Т. 89. № 1. С. 99–110. DOI: 10.1134/S0026365620010152.
- Gohl D.M., MacLean A., Hauge A. et al. An optimized protocol for high-throughput amplicon-based microbiome profiling // Research Square. 2016. Nо. 1. Art. 30. DOI: 10.1038/protex.2016.030.
- Fadrosh D.W., Ma B., Gajer P. et al. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform // Microbiome. 2014. Nо. 2 (1). Art. 6. DOI: 10.1186/2049-2618-2-6.
- Hugerth L.W., Wefer H.A., Lundin S. et al. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies // Applied and Environmental Microbiology. 2014. Nо. 80 (16). P. 5116–5123. DOI: 10.1128/AEM.01403-14.
- Merkel A.Y., Podosokorskaya O.A., Chernyh N.A., Bonch Osmolovskaya E.A. Occurrence, diversity, and abundance of methanogenic archaea in terrestrial hot springs of Kamchatka and Sao Miguel Island // Microbiology. 2015. Vol. 84. P. 577–583.
- Srikanth M., Sandeep T.S.R.S., Sucharitha K., Godi S. Biodegradation of plastic polymers by fungi: a brief review // Bioresources Bioprocessing. 2022. Vol. 9. Art. 42. DOI: 10.1186/s40643-022-00532-4.
- Kanamaru K., Kashiwagi S., Mizuno T. A copper-transporting P-type ATPase found in the thylakoid membrane of the cyanobacterium Synechococcus species PCC7942 // Molecular Microbiology. 1994. Vol. 13 (2). P. 369–377. DOI: 10.1111/j.1365-2958.1994.tb00430.x.
- Kanehisa M., Goto S. KEGG: Kyoto encyclopedia of genes and genomes // Nucleic Acids Research. 2000. Vol. 28. P. 27–30. DOI: 10.1093/nar/28.1.27.
- Абдуллин И.Ш., Канарская З.А., Хубатхузин А.А. и др. Нанодисперсные материалы на основе титана в микробиологической, медицинской и пищевой промышленностях // Вестник Казанского технологического университета. 2012. Т. 15. № 11. С. 158–165.
- Bonyadi Z., Mirzaee M., Ejtehadi M.M., Mokhtari M. The bactericidal effect of simultaneous titanium oxide on common hospital bacteria // Environmental Monitoring and Assessment. 2017. Vol. 189. Art. 342. DOI: 10.1007/s10661-017-6049-5.
- Nies D.H. Efflux-mediated heavy metal resistance in prokaryotes // Microbiology Review. 2003. Vol. 27 (2-3). P. 313–339. DOI: 10.1016/S0168-6445(03)00048-2.
- Choudhury R., Srivastava S. Zinc resistance mechanisms in bacteria // Current Science. 2001. Vol. 81. P. 768–775.
- Viti C., Marchi E., Decorosi F., Giovannetti L. Molecular mechanisms of Cr(VI) resistance in bacteria and fungi // Microbiology Review. 2014. Vol. 38 (4). P. 633–659. DOI: 10.1111/1574-6976.12051.
- Hynninen A., Touzé T., Pitkänen L. et al. An efflux transporter PbrA and a phosphatase PbrB cooperate in a lead-resistance mechanism in bacteria // Molecular Microbiology. 2009. Vol. 74 (2). P. 384–394. DOI: 10.1111/j.1365-2958.2009.06868.x.
- Воробьев А.В. Полиэфирные смолы // Компоненты и технологии. 2003. № 6. С. 182–185.