Идентификация коллекции мицелиальных грибов НИЦ «Курчатовский институт» – ВИАМ молекулярно-генетическими методами
Проведена таксономическая идентификация штаммов мицелиальных грибов коллекции НИЦ «Курчатовский институт» – ВИАМ с помощью молекулярно-генетических методов. Таксономическая классификация проведена в Курчатовском геномном центре с помощью двух подходов: анализ геномных сигнатур и полноценный филогенетический анализ по 255 маркерным однокопийным генам эукариот. Представлены данные молекулярной идентификации штаммов следующих таксонов: род Penicillium; порядок Xylariales, роды Nigrospora, Neopestalotiopsis, Pestalotiopsis, Daldinia; род Cladosporium; род Sarocladium; порядок Hypocreales; порядок Mucorales, род Rhizopus; род Trichoderma; род Fusarium и др.
Введение
В микробиологической лаборатории НИЦ «Курчатовский институт» – ВИАМ ведутся испытания и исследования устойчивости материалов к микроорганизмам. В рамках этой работы сотрудники лаборатории постоянно выделяют и идентифицируют микрофлору, поражающую технику и конструкции. За эти годы сформирована обширная коллекция микроорганизмов, способных разрушать материалы [1–3]. Преимущественно она состоит из культур микроскопических грибов, выделенных из образцов, прошедших испытания в различных климатических условиях. Кроме того, в лаборатории анализируется техника с признаками микробиологических повреждений, что позволяет получать культуры деструкторов непосредственно из очагов поражения. Эти штаммы особенно ценны для дальнейших исследований их свойств и тестирования материалов на устойчивость [4–8].
Ранее идентификация микроорганизмов основывалась на морфологических признаках. Такой подход был ненадежным из-за изменчивости внешних характеристик и ограниченности данных микробиологических справочников. С появлением методов секвенирования точность определения микроорганизмов значительно возросла и позволяет идентифицировать культуры на видовом уровне.
Цель данной работы – таксономическая идентификация культур мицелиальных грибов депозитария НИЦ «Курчатовский институт» – ВИАМ с помощью методов секвенирования.
Материалы и методы
На первом этапе проведена реизоляция 200 культур мицелиальных грибов из замороженных и лиофилизированных препаратов, хранящихся в депозитарии НИЦ «Курчатовский институт» – ВИАМ, путем высева инокулюма на поверхность питательной среды сусло-агар. Чашки Петри с инокулюмом заклеивали парафильмом и инкубировали в термостатах при температуре 28 °С в течение 10–14 дней. После проведения инкубации проверяли идентичность и чистоту культур, при необходимости повторяли процедуру пересевов на новые чашки Петри с питательной средой.
Молекулярную диагностику проводили в Курчатовском геномном центре с помощью двух подходов: анализ геномных сигнатур и полноценный филогенетический анализ по 255 маркерным однокопийным генам эукариот.
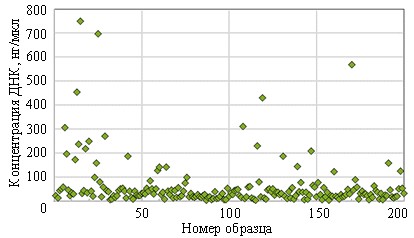
Сначала проводили выделение геномной ДНК с культур на чашках Петри (см. первую часть данной статьи). Далее концентрацию полученной ДНК измеряли на флуориметре. В ходе измерения в образец вводится флуоресцентный краситель, который специфически связывается с целевой молекулой ДНК, образуя флуоресцентный комплекс. Прибор измеряет интенсивность свечения и рассчитывает концентрацию относительно известных стандартов. В результате измерения концентраций получены удовлетворительные результаты. Сводные результаты представлены на рис. 1.
Качество препаратов ДНК определяли на спектрофотометре. По полученному графику спектра поглощения проверяли наличие примесей в препарате ДНК. Так, по соотношению значений поглощения при длинах волн 260 и 280 нм оценивали наличие примесей белков и других соединений с пиком поглощения в области 280 нм. В свою очередь, по соотношению значений поглощения при длинах волн 260 и 230 нм определяли наличие примесей солей гуанидина (присутствует в лидирующем растворе наборов реагентов для экстракции ДНК) и других соединений с пиком поглощения в области 230 нм. Пример графика спектра поглощения представлен на рис. 2.
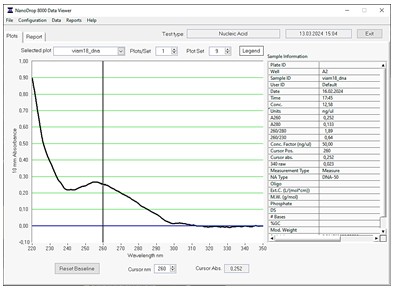
Далее проводили дополнительную очистку полученных препаратов ДНК. Несмотря на наличие достаточного количества ДНК для приготовления геномных библиотек, в ряде образцов наблюдали низкое качество полученного материала. Если соотношение значений поглощения при длинах волн 260 и 280 нм приближалось к оптимальному (~(1,8±0,2)), то при длинах волн 260 и 230 нм в большинстве образцов было существенно меньше необходимого (2,0–2,2). Низкое качество исходной ДНК может привести к некорректной работе реагентов для приготовления библиотек, поэтому для каждого образца проведена дополнительная очистка 500 нг ДНК с помощью набора реагентов для очистки ДНК.
Качество ДНК определяли на спектрофотометре. По полученному графику спектра поглощения проверяли наличие остаточных примесей в препарате ДНК. В примере, приведенном на рис. 3, демонстрируется значительное повышение качества препарата ДНК штамма viam 018 по сравнению с тем же препаратом без дополнительной очистки (рис. 2).
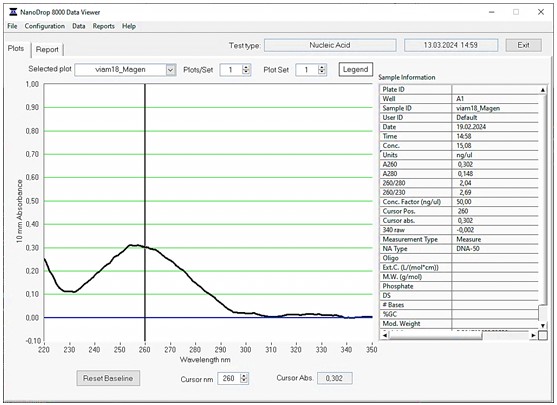
Далее для очищенной ДНК культур на чашках Петри приготовлены библиотеки для секвенирования, проведена оценка качества и количества полученных библиотек. После осуществлена биоинформативная обработка данных, включающая предварительную обработку и контроль качества геномных прочтений и de novo сборку геномов.
Работа выполнена при поддержке ЦКП «Климатические испытания» НИЦ «Курчатовский институт» – ВИАМ.
Результаты и обсуждение
В рамках данной работы с помощью анализа NGS-данных (Next Generation Sequencing – секвенирование нового поколения) решали две основные задачи: точная таксономическая идентификация 200 штаммов мицелиальных грибов депозитария НИЦ «Курчатовский институт» – ВИАМ и de novo сборка и структурно-функциональная аннотация геномов. Подходы к решению второй задачи осуществляются с помощью стандартных биоинформатических инструментов, если исследуемые культуры являются чистыми монокультурами. В то же время даже небольшое количество контаминантов в культуре может приводить к значительному ухудшению качества сборки и, следовательно, результатов анализа. Культуры мицелиальных грибов нередко оказываются загрязнены сопутствующими бактериальными микроорганизмами, поэтому в рамках данного проекта использовали нормализацию ридов по k-мерам с помощью пакета BBDuk, позволяющую отсечь незначительную (до 10 %) контаминацию [9]. Однако, несмотря на это, ряд образцов все же оказался сильно загрязнен или не представлял собой монокультуру.
Точное определение таксономии грибов не является тривиальной задачей в первую очередь по причине того, что, в отличие от прокариотических организмов [10], универсальных инструментов для этих целей не разработано. По этой причине таксономию определяли с помощью мультифазного подхода. На первом этапе проводили поиск геномных сигнатур, позволяющих с высокой точностью идентифицировать те штаммы, для которых есть геномы близкородственных организмов (уровень вида/подвида) в референсных базах данных. Однако использование данного подхода, во-первых, требует подтверждения путем проведения полноценного филогенетического анализа, во-вторых, не эффективно в случаях, когда информация по геномам близкородственных организмов отсутствует. Для решения этой проблемы проводили филогенетический анализ на основании последовательностей маркерных генов и строили филогенетическое древо, включающее как исследуемые образцы, так и геномы типовых штаммов грибов, доступных в открытых источниках.
На заключительном этапе проведена структурная и функциональная аннотация геномов для получения данных о представленности различных категорий кластеров ортологичных генов и последовательностей генов рРНК [11–14].
Общая схема анализа представлена на рис. 4.
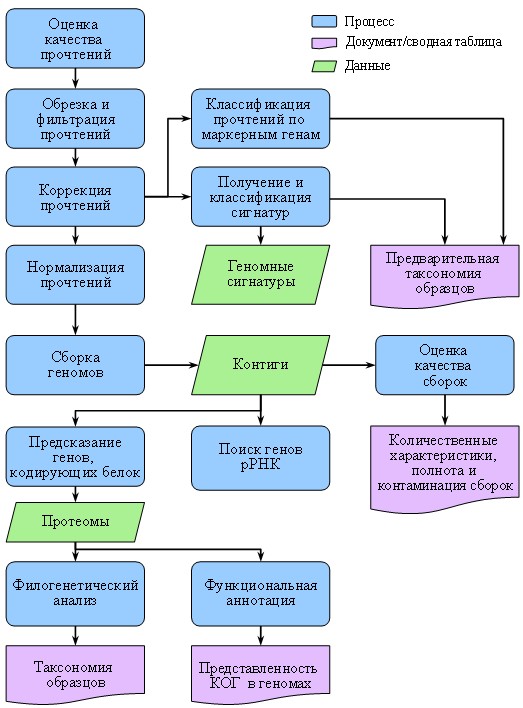
В предыдущей части данной статьи представлены данные по молекулярной идентификации штаммов следующих таксонов: порядок Glomerellales, род Colletotrichum; семейство Pleosporaceae; роды Curvularia и Alternaria, род Aspergillus и представители новых порядков класса Sordariomycetes.
С помощью анализа геномных сигнатур [15–18] к представителям рода Penicillium отнесены штаммы viam 017, viam 019, viam 049, viam 072, viam 077, viam 082, viam 084, viam 085, viam 086-1, viam 086-2, viam 087, viam 088, viam 089, viam 093, viam 094-1, viam 094-2, viam 104, viam 106, viam 107, viam 129, viam 135, viam 150, viam 151, viam 164, viam 168, viam 185, viam 186, viam 187, viam 231 и viam 234. С помощью филогенетического анализа подтверждена филогения всех этих штаммов и классифицирован ряд штаммов, для которых метод анализа геномных сигнатур оказался недостаточно чувствительным (рис. 5).
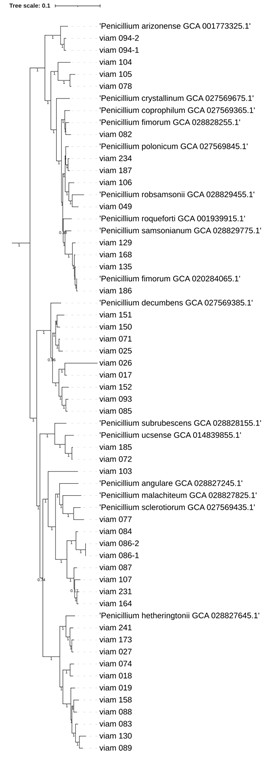
Анализ филогенетического древа показывает, что в большинстве случаев исследуемые штаммы оказываются близки типовым видам рода Penicillium, однако окончательное подтверждение их видовой принадлежности требует более глубокого мультифазного подхода с использованием методов анализа средней нуклеотидной/аминокислотной идентичности, ключевых метаболических путей и др. В то же время ряд штаммов, например viam 104, viam 103, а также кластеры viam 151/
viam 150/viam 071/viam 025, viam 087/viam 107/viam 231/viam 164 и др. (рис. 5), вероятно, представляют собой новые виды рода Penicillium.
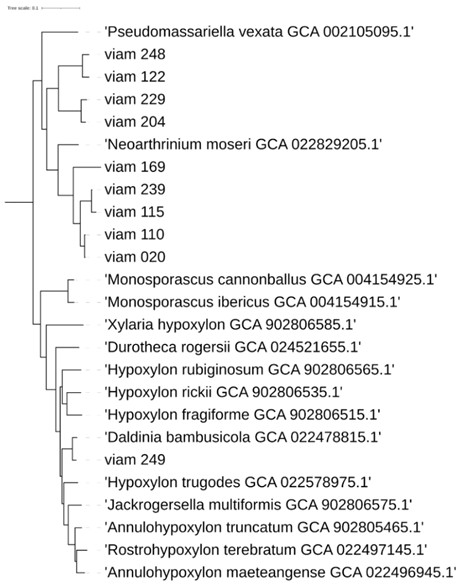
С помощью анализа геномных сигнатур к представителям порядка Xylariales отнесены штаммы viam 020, viam 110, viam 115, viam 122, viam 204, viam 229, viam 239, viam 248 и viam 249.
Филогенетический анализ подтвердил принадлежность этих штаммов к данному порядку, к которому также отнесен штамм viam 169 (рис. 6). В то же время в результате отсутствия ряда геномов типовых штаммов в базе данных в некоторых случаях классификация с помощью анализа геномных сигнатур была более точной. Так, штаммы viam 248 и viam 122 отнесены к роду Nigrospora; штаммы viam 115 и viam 239 – к роду Neopestalotiopsis; штамм viam 020 (и, судя по филогенетическому древу, штамм viam 110) – к роду Pestalotiopsis. Штамм viam 249, по-видимому, относится к виду Daldinia bambusicola.
Следует отметить, что, судя по глубине ветвления, штамм viam 169 представляет собой новый род семейства Apiosporaceae, к которому относится род Neoarthrinium.
С помощью анализа геномных сигнатур к представителям рода Cladosporium отнесены штаммы viam 096, viam 109 cl3, viam 118, viam 134, viam 145, viam 148, viam 157, viam 182, viam 210 и viam 223. С помощью филогенетического анализа подтверждена филогения всех этих штаммов, а также классифицировано значительное количество штаммов, для которых метод анализа геномных сигнатур оказался недостаточно чувствительным (рис. 7).
Анализ филогенетического древа показывает, что все штаммы этой ветви близки единственному типовому штамму рода Cladosporium (согласно базе данных геномов типовых штаммов), однако окончательное подтверждение их видовой принадлежности требует глубокого мультифазного подхода с использованием методов анализа средней нуклеотидной/аминокислотной идентичности и расширенных баз данных по мере появления новых геномов рода Cladosporium.

Далее проведена идентификация штаммов viam 240, viam 029, viam 099 и viam 076 cl3. Анализ геномных сигнатур показал, что штамм viam 240 относится к виду Sarocladium implicatum, viam 029 – к виду Golovinomyces magnicellulatus, а штаммы viam 099 и viam 076 cl3 – к роду Clonostachys.
Данные филогенетического анализа (рис. 8) подтверждают принадлежность штамма viam 240 к роду Sarocladium, но к другому виду – Sarocladium brachiariae. Штамм viam 029 кластеризовался с видом Emericellopsis atlantica, который, как и G. magnicellulatus, принадлежит к отделу Ascomycota, но к другому классу – Leotiomycetes. Штаммы viam 099 и viam 076 cl3 образуют единую монофилетическую группу, кластеризующуюся с видом Gliomastix tumulicola, принадлежащим с родом Clonostachys к одному семейству Bionectriaceae, порядка Hypocreales.

Анализ геномных сигнатур показал, что штамм viam 037 относится к роду Rhizopus, что подтверждается результатами филогенетического анализа (рис. 9).

С помощью анализа геномных сигнатур к представителям рода Trichoderma отнесены штаммы viam 160, viam 073, viam 051, viam 108, viam 112, viam 116, viam 050, viam 038, viam 176, viam 166, viam 136 и viam 216. Анализ филогенетического древа подтверждает принадлежность к этому роду Trichoderma также штаммов viam 153 и viam 069 (рис. 10). Более того, штамм viam 216 ближе к виду Trichoderma brevicrassum, viam 038 – к виду Trichoderma harzianum, а viam 108, viam 112 и viam 116 – к виду Trichoderma asperellum.

Анализ данных по геномным сигнатурам показал, что штаммы viam 060, viam 225, viam 121, viam 128, viam 212, viam 183, viam 246, viam 068, viam 039 и viam 070 относятся к представителям рода Fusarium. Анализ филогенетического древа подтверждает принадлежность данных штаммов к роду Fusarium (рис. 11). При этом видно, что штамм viam 070 ближе всего к виду Fusarium poae, штамм viam 039 – к виду Fusarium goolgardi. Штаммы viam 225 и viam 060 кластеризуются с видом Fusarium tricinctum, однако, вероятно, это представители другого вида. Штаммы viam 121, viam 128, viam 212, viam 183, viam 246 и viam 068, по-видимому, являются новыми видами рода Fusarium.

Заключения
Проведена таксономическая идентификация штаммов мицелиальных грибов коллекции НИЦ «Курчатовский институт» – ВИАМ с помощью молекулярно-генетических методов. Работа по проведению полногеномного секвенирования выполнена в Курчатовском геномном центре.
На первом этапе осуществлена молекулярная идентификация штаммов следующих таксонов: порядок Glomerellales, род Colletotrichum; семейство Pleosporaceae; роды Curvularia и Alternaria, род Aspergillus и представители новых порядков класса Sordariomycetes. В данной части статьи представлены результаты молекулярной идентификации штаммов следующих таксонов: род Penicillium; порядок Xylariales, роды Nigrospora, Neopestalotiopsis, Pestalotiopsis, Daldinia; род Cladosporium; род Sarocladium; порядок Hypocreales; порядок Mucorales, род Rhizopus; род Trichoderma; род Fusarium и др. Заключительный анализ данных по идентификации других штаммов будет представлен в следующей публикации.
- Каблов Е.Н., Лаптев А.Б., Прокопенко А.Н., Гуляев А.И. Релаксация полимерных композиционных материалов под длительным действием статической нагрузки и климата (обзор). Часть 1. Связующие // Авиационные материалы и технологии. 2021. № 4 (65). С. 70–80. URL: http://www.journal.viam.ru (дата обращения: 20.07.2025). DOI: 10.18577/2713-0193-2021-0-4-70-80.
- Закирова Л.И., Афанасьев-Ходыкин А.Н., Мовенко Д.А., Лаптев А.Б. Особенности формирования диффузионного слоя Sn–Zn–Fe на границе гальванотермического покрытия системы цинк–олово и стали 30ХГСА с высокой защитной способностью // Авиационные материалы и технологии. 2022. № 4 (69). С. 61–71. URL: http://www.journal.viam.ru (дата обращения: 29.07.2025). DOI: 10.18577/2713-0193-2022-0-4-61-71.
- Старцев В.О., Старцев О.В., Зеленева Т.О., Варданян А.М. Влияние осадков на изменение массы образцов полимерных композиционных материалов в открытых климатических условиях // Авиационные материалы и технологии. 2024. № 1 (74). С. 136–154. URL: https://www.journal.viam.ru (дата обращения: 20.07.2025). DOI: 10.18577/2713-0193-2024-0-1-136-154.
- Lugauskas A., Levinskait L., Pečiulyt D. Micromycetes as deterioration agents of polymeric materials // International biodeterioration & biodegradation. 2003. Vol. 52 (4). P. 233–242.
- Lugauskas A., Prosychevas I., Levinskaitė L., Jaskelevičius B. Physical and chemical aspects of long‐term biodeterioration of some polymers and composites // Environmental Toxicology: An International Journal. 2004. Vol. 19 (4). P. 318–328.
- Srikanth M., Sandeep T.S.R.S., Sucharitha K., Godi S. Biodegradation of plastic polymers by fungi: a brief review // Bioresources and Bioprocessing. 2022. Vol. 9 (1). P. 42. DOI: 10.1186/s40643-022-00532-4.
- Кривушина А.А., Лаптев А.Б. Фунгициды: применение, свойства и принципы действия // Авиационные материалы и технологии. 2024. № 1 (74). С. 155–168. URL: http://www.journal.viam.ru (дата обращения: 20.07.2025). DOI: 10.18577/2713-0193-2024-0-1-155-168.
- Каблов Е.Н., Антипов В.В. Роль материалов нового поколения в обеспечении технологического суверенитета Российской Федерации // Вестник Российской академии наук. 2023. Т. 93. № 10. С. 907–916. DOI: 10.31857/S0869587323100055.
- Bushnell B. BBMap: A Fast, Accurate, Splice-Aware Aligner. LBL publications, 2014. 5 р.
- Chaumeil P.A., Mussig A.J., Hugenholtz P. et al. GTDB-Tk: a toolkit to classify genomes with the Genome Taxonomy Database. Oxford University Press, 2020. P. 1925–1927.
- Jeon S.A., Park J.L., Park S.J. et al. Comparison between MGI and Illumina sequencing platforms for whole genome sequencing // Genes and Genomics. 2021. Vol. 43. No. 7. P. 713–724. DOI: 10.1007/s13258-021-01096-x.
- Hu T., Che J., Lin X. et al. Comparison of the DNBSEQ platform and Illumina HiSeq 2000 for bacterial genome assembly // Scientific Reports. 2024. Vol. 14. No. 1. P. 1292. DOI: 10.1038/s41598-024-51725-0.
- Meslier V., Quinquis B., Da Silva K. et al. Benchmarking second and third-generation sequencing platforms for microbial metagenomics: 1 // Scientific Data. 2022. Vol. 9. No. 1. P. 694.
- Blanco-Míguez A., Beghini F., Cumbo F. et al. Extending and improving metagenomic taxonomic profiling with uncharacterized species using MetaPhlAn 4 // Nature biotechnology. Nature Publishing Group, 2023. P. 1–12.
- Manni M., Berkele M.R., Seppey M. et al. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes // Molecular biology and evolution. 2021. Vol. 38. No. 10. P. 4647–4654.
- Katoh K., Standley D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability // Molecular biology and evolution. 2013. Vol. 30. No. 4. P. 772–780.
- Capella-Gutiérrez S., Silla-Martínez J.M., Gabaldón T. trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses // Bioinformatics. 2009. Vol. 25. No. 15. P. 1972–1973.
- Price M.N., Dehal P.S., Arkin A.P. FastTree 2 – approximately maximum-likelihood trees for large alignments // PLoS One. 2010. Vol. 5. No. 3. P. e9490.