Выделение бактериальных штаммов деструкторов полимерных материалов из образцов оборотной охлаждающей воды нефтеперерабатывающего и нефтехимического заводов
Представлены методы выделения бактериальных штаммов потенциальных деструкторов полимерных материалов из образцов оборотной воды предприятий по производству нефтепродуктов. Культуры экспонировали с полимерными материалами и полимерными субстратами. Данные о видовом составе бактериальных культур получены в результате секвенирования ДНК-фрагмента, кодирующего V4-участок 16S рРНК бактерий. Проведена сравнительная оценка кинетики роста чистых культур бактерий рода Pseudomonas в минеральной среде с добавлением реактивного и дизельного топлив.
Введение
Первичные продукты нефтепереработки применяются для синтеза исходных компонентов в производстве полимерных материалов. Полиэфирные и поливиниловые волокна, адгезивные покрытия, связующие и клеи в составе полимерных композиционных материалов могут иметь множество вариантов исполнения в зависимости от назначения конечного продукта. Такие полимерные материалы, как полиэтилентерефталат (ПЭТФ) и поливинилхлорид, полиакрилаты и эпоксидные смолы, широко распространены благодаря уникальным физико-химическим свойствам, механическим характеристикам и стойкости к воздействию окружающей среды.
Учитывая совокупный объем произведенных и импортированных производных олефиновой и ароматической фракций нефти, насчитывающих десятки наименований, можно предположить, что как минимум несколько сотен тысяч тонн полимерных материалов не будут подвергнуты рециклингу или полной переработке и останутся в окружающей среде. Это обстоятельство является серьезной экологической проблемой, поскольку ни один из упомянутых полимерных материалов не подвержен быстрому биологическому разложению [1, 2].
Скорость процесса биодеструкции полимеров в окружающей среде в первую очередь определяется химическим составом и степенью кристалличности материала, а также интенсивностью воздействия климатических факторов, важнейшими из которых являются температура и влажность [3–8]. Происходит изменение физико-механических (потеря формы, массы, снижение вязкости, гибкости, охрупчивание) и химических (разрыв химических связей, снижение молекулярной массы мономеров) свойств материала. На этапе дезинтеграции материал получает необратимые повреждения, которые при участии микроорганизмов сначала приводят к фрагментации материала и в дальнейшем к его минерализации, подразумевающей полную деполимеризацию с образованием диоксида углерода, метана и воды в зависимости от конечного акцептора электронов [9–12].
Одним из наиболее часто упоминаемых бактериальных штаммов биодеструкторов является Ideonella sakaiensis 201-F6, для которого установлен биохимический механизм утилизации ПЭТФ, включая структуру и активность ключевых ферментов деполимеризации и дальнейшей метаболической ассимиляции частей мономера – терефталевой кислоты и этиленгликоля [13, 14]. С химической точки зрения ПЭТФ является инертным полимером за счет наличия ароматических компонентов с эфирными связями в составе мономера. Однако ПЭТФ, использованный в эксперименте с культурой Ideonella sakaiensis 201-F6, имел достаточно низкую кристалличность (1,9 %), в то время как ПЭТФ, применяемый для изготовления тары и упаковок, имеет кристалличность от 30 до 40 %, что существенно снижает его биодоступность [1, 13, 14].
В данной работе демонстрируется практическая возможность получения из сложных эндемических сообществ микроорганизмов чистых культур бактериальных штаммов рода Pseudomonas как наиболее активных в отношении органических соединений, в том числе токсичных, и растворимых в воде полимеров, применяемых в производстве материалов.
Материалы и методы
Химический состав оборотной охлаждающей воды нефтеперерабатывающего (НПЗ) и нефтехимического (НХЗ) заводов определен в испытательном лабораторном центре ФБУЗ «Центр гигиены и эпидемиологии в республике Башкортостан» в соответствии с сертифицированными методами исследования (см. приложение, табл. 1 и 2).
Для экспонирования с бактериальной культурой использовали образцы материалов размером 25×70×1,5 мм (табл. 1). Перед взаимодействием с микроорганизмами поверхность контрольных и опытных образцов, размещенных на пластиковой подложке, обрабатывали раствором 0,3%-ной соляной кислоты и выдерживали в течение 12 ч при комнатной температуре. Затем дважды образцы выдерживали в течение 6 ч в камере светопогоды в диапазоне длин волн ультрафиолетового излучения 300–400 нм при плотности потока 65 Вт/м2 и относительной влажности ~25 %.
Таблица 1
Материалы, использованные для экспонирования с бактериальной культурой
Образцы материалов фотографировали с помощью зеркальной фотокамеры Canon EOS 5D Mark III с объективом EF 100mm f/2.8L Macro.
Для получения первичных бактериальных культур из каждого образца воды НПЗ и НХЗ в соответствии с общим микробным числом (ОМЧ) отбирали определенный объем и переносили в 50 мл базовой минеральной среды с добавлением 1 % (объемн.) стерильного дизельного топлива марки ДТ-Л-40-К2 (ГОСТ 305–2013). Состав среды, мМ: 7 – KH2PO4; 5 – K2HPO4; 20 – NH4Cl; 10 – NaCl; 0,8 – MgSO4·7H2O; 0,18 – CaCl2; pH = 7.
Базовую среду стерилизовали автоклавированием при температуре 121 °C в течение 15 мин.
Для замедления или предотвращения возможного роста микромицетов добавляли 50 мкл раствора амфотерицина В в 50%-ном этиловом спирте с исходной концентрацией действующего вещества 100 мг/мл.
Культуры наращивали в течение не менее 72 ч при температуре 25 °C и скорости перемешивания 140 мин–1.
Полученные первичные бактериальные культуры высевали на твердую базовую минеральную среду, содержащую 1 % агарозы и 0,01 % дрожжевого экстракта, для проведения полимеразной цепной реакции (ПЦР) отдельных колоний и идентификации секвенированием.
Для экспонирования материалов подготовленные опытные образцы помещали в 20 мл базовой минеральной среды, содержащей смесь первичных бактериальных культур плотностью ~0,05 ОЕ/мл, полученную смешиванием в равных пропорциях первичных культур для каждого образца воды НПЗ и НХЗ. Контрольные образцы материалов помещали в 20 мл базовой минеральной среды без добавления бактериальных культур. Контрольные и опытные образцы материалов экспонировали в течение 60 дней при температуре 25 °C и скорости перемешивания 50 мин–1. В табл. 2 приведены сведения о соответствии наименований образцов материалов и полученных проб для секвенирования.
Этап отбора | Пробы для секвенирования после экспонирования с материалом | ||||
дизельное ДТ-Л-40-К2 | клеевая пленка ВК-36 | компонент А | компонент Б | стирол-акриловая | |
1 | D-МС | – | – | – | – |
2 | – | AG-Mix | |||
3 | – | A2 PVA | E1 PVA | F2 PVA | C3 PVA |
4 | – | – | E1 SA | F2 SA | – |
| Примечание. D-MC – проба первичной культуры; AG-Mix – проба после экспонирования первичной культуры с образцами материалов; PVA – поливиниловый спирт; SA – стирол-акриловая суспензия. | |||||
Для получения чистых культур штаммов бактерий проводили рассев аликвот культуральной жидкости, полученной после экспонирования первичных культур с материалами, на твердые базовые минеральные среды, содержащие 1 % агарозы и 0,025 % стерильных субстратов акриловой дисперсии или поливинилового спирта. Полученные отдельные колонии снова засевали в жидкие базовые минеральные среды с теми же субстратами. Полученные отдельные колонии идентифицировали секвенированием.
При подсчете колоний выявлено низкое ОМЧ при экспонировании с клеевой пленкой ВК-102А (B2), поэтому в целях экономии расходных материалов и времени данная проба исключена из дальнейшего отбора.
Для получения данных о показателях роста выделенных чистых культур бактерий на топливах использовали 3 мл базовой минеральной среды с добавлением 3 мл дизельного топлива марки ДТ-Л-40-К2 (ГОСТ 305–2013) или реактивного топлива марки ТС-1 (ГОСТ 10227–86). Данные о кинетике роста получали в течение 96 ч при температуре 25 °C и начальной плотности исследуемых культур 0,025 ОЕ/мл без перемешивания, пробирки в течение всего времени испытаний находились в вертикальном положении. В качестве отрицательного контроля использовали чистую среду с указанными видами топлив.
Оптическую плотность бактериальных культур в контрольных и опытных образцах определяли с помощью спектрофотомера ПЭ-5000 (Экрос, Россия) при длине волны 600 нм. Кинетику роста определяли по формуле
K = tg(A24 – A0),
где А0 и А24 – поглощение среды исходное и через 24 ч при длине волны 600 нм.
Для амплификации V4-фрагментов 16S рРНК бактерий использовали модифицированные олигонуклеотидные праймеры 515f и 806r [15]. Концентрацию препаратов ДНК определяли флюориметрическим методом с помощью набора QuDye BR (Люмекс, Россия) согласно инструкции. Геномную ДНК, полученную с помощью набора для выделения ДНК на селикагелевых колонках, в количестве ~50 нг добавляли в 100 мкл смеси для ПЦР, содержащей HS Taq ДНК-полимеразу, реакционный буфер, дезоксинуклеозидтрифосфаты и соответствующие синтетические олигонуклеотиды (Евроген, Россия). Реакции выполняли в автоматическом цифровом программируемом термоциклере (амплификаторе) с нагреваемой крышкой при следующих параметрах: 95 °C в течение 5 мин; 95 °C в течение 30 с, 60 °C в течение 30 с, 72 °C в течение 10 с (30 циклов); 72 °C в течение 5 мин. Полученные ПЦР-фрагменты очищали с помощью реагента фракционирования ДНК на магнитных частицах в соответствии с инструкцией производителя.
Результаты ПЦР визуализировали с помощью электрофореза в 1,5%-ном агарозном геле для молекулярной биологии в однократном трис-ацетат-ЭДТА буфере с красителем бромид этидия в количестве 0,5 мкг/мл (Хеликон, Россия). Для электрофореза использовали четырехканальный источник питания с выходным напряжением 5000 В с постоянным током 8 В/см агарозного геля. Для определения размера ДНК-фрагментов, получаемых в результате ПЦР, использовали маркеры молекулярной массы с размерами 100, 200, 300, 500, 700, 1000, 1500, 2000, 3000, 5000, 7000 и 10000 пар нуклеотидов.
Для амплификации ДНК полноразмерного фрагмента 16S рРНК бактерий использовали модифицированные олигонуклеотиды 27f и 1492r [16]. Параметры реакции: 95 °C в течение 5 мин; 95 °C в течение 30 с, 58 °C в течение 30 с, 72 °C в течение 1 мин (30 циклов); 72 °C в течение 5 мин. Для выделения ДНК из агарозного геля использовали набор LumiPure (Lumiprobe, Россия). Результаты ПЦР визуализировали с помощью электрофореза с указанными ранее параметрами и оборудованием. Данные о первичной структуре выделенных ДНК-фрагментов получены в компании Евроген (Москва) на генетическом анализаторе 3500хL. Сборка и валидация перекрывающихся прочтений выполнены в специализированной программе [17].
Данные коротких прочтений V4-фрагмента гена 16S рРНК бактерий получены с помощью высокопроизводительного секвенатора с набором реагентов для секвенирования nano kit, v2 (500 cycles), работы выполнены согласно официальному руководству компании изготовителя. Качество прочтений анализировали с помощью соответствующей программы (см. приложение, рис. 1) [18]. Удаление адаптеров, сборка перекрывающихся прочтений, выравнивание по базе данных первичных структур гена 16S рРНК архей и бактерий, визуализации выполнены с применением модулей программы reporter v.2.6.2.3.
Для ПЦР-исследования отдельных колоний первичных культур использовали специфичные для рода Pseudomonas праймеры PSF2 и PAGS-R [19]. Реакции выполняли при следующих параметрах: 95 °C в течение 3 мин; 95 °C в течение 30 с, 60 °C в течение 30 с, 72 °C в течение 30 с (30 циклов); 72 °C в течение 5 мин. Для выявления наличия плазмид incP9 использовали олигонуклеотиды rep9F и rep9R [20]. Для поиска генов диоксигеназ ароматических соединений применяли праймеры TOD-F и TOD-R [21]. Реакции выполняли при следующих параметрах: 95 °C в течение 3 мин; 95 °C в течение 30 с, 55 °C в течение 30 с, 72 °C в течение 30 с (35 циклов); 72 °C в течение 5 мин. Реагенты и оборудование для ПЦР указаны ранее. Результаты ПЦР визуализировали с помощью электрофореза с указанными ранее параметрами и оборудованием.
Работа выполнена при поддержке ЦКП «Климатические испытания» НИЦ «Курчатовский институт» – ВИАМ.
Результаты и обсуждение
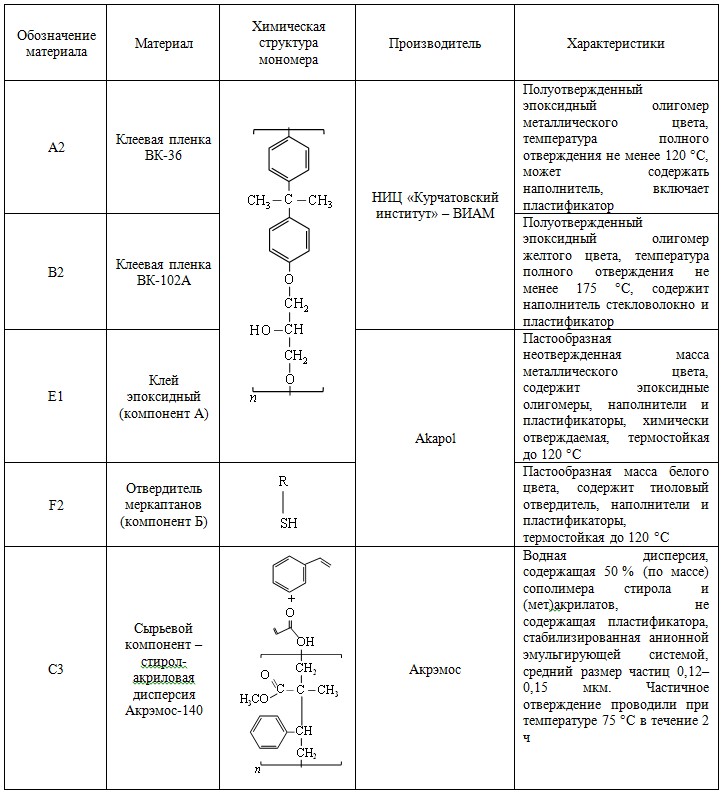
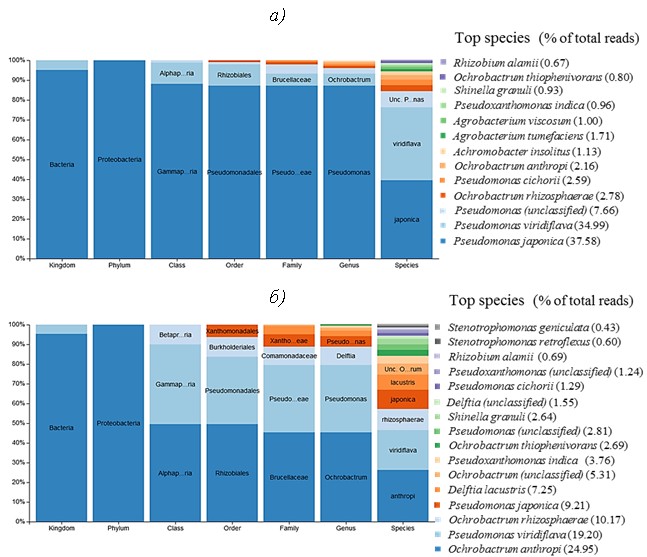
Таксономическая идентификация первичных бактериальных культур, полученных на первом этапе отбора, демонстрирует, что образец на 95 % состоит из бактерий рода Pseudomonas (рис. 1). Такой результат связан в первую очередь с составом базовой минеральной среды и, по-видимому, с высоким содержанием бактерий указанного рода в исходных образцах оборотной охлаждающей воды НПЗ и НХЗ.
На этом этапе решено провести ПЦР-исследование отдельных колоний первичных культур для выяснения эффективности такого подхода к поиску целевых фенотипов бактериальных штаммов. Для этого выбраны две последовательности мишени, одна из которых представляет собой участок плазмидного репликонаincP9, а вторая – участок гена todC1,кодирующего диоксигеназу бензола, толуола и хлорбензола. При этом ген todC1, по-видимому, имеет хромосомную локализацию в бактериальной клетке.
С учетом того, что указанные плазмиды и ген todC1 найдены исключительно в бактериях рода Pseudomonas, необходимо установить, что каждая из отобранных колоний относится к этому роду (см. приложение, рис. 2).
Как можно убедиться, каждая из 18 отобранных колоний является представителем рода Pseudomonas. Эти данные хорошо согласуются с анализом 16S рРНК первичных культур в пробе D-MC, которые служили источником для ПЦР-исследования.
Дальнейший ПЦР-анализ 18 образцов с олигонуклеотидами rep9F/rep9R и TOD-F/TOD-R выявил предположительное наличие этих последовательностей ДНК только в одном образце (см. приложение, рис. 3).
Хотя для полной уверенности в том, что амплифицированные фрагменты ДНК действительно являются участками repA и todC1, необходимо секвенирование полученных в результате ПЦР фрагментов ДНК. Данный образец культуры выбран для дальнейшего анализа в качестве результата применения такого подхода к поиску редких фенотипов бактерий.
Полученный штамм в одном из запусков секвенирования в качестве отдельного образца идентифицирован по V4-фрагменту 16S рРНК как Pseudomonas plecoglossicida (см. приложение, рис. 4).
Перед экспонированием с первичной культурой образцы материалов целенаправленно повреждали с помощью соляной кислоты и УФ-излучения (см. приложение, рис. 5). Одним из результатов такого рода воздействий является также полная стерильность каждого образца материала вследствие длительного воздействия перечисленных факторов.
Для экспонирования с первичной культурой бактерий не использовали контрольные образцы материалов, не прошедшие подготовку описанным способом. Это связано с тем, что с учетом указанной цели исследования испытаниям подвергали культуру микроорганизмов, а материалы служили в качестве инструмента, поэтому отрицательными контролями выступили прошедшие соответствующую подготовку материалы без добавления к стерильной среде микроорганизмов.
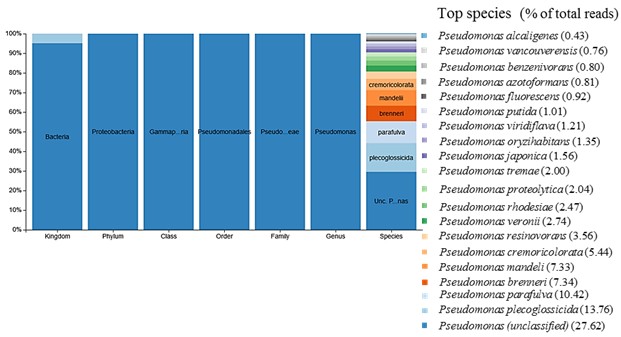
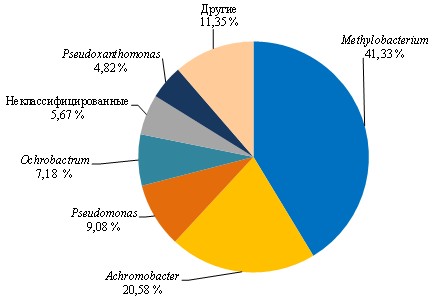
После 60 дней экспонирования из каждого образца материалов с первичной культурой бактерий отобрана часть среды для получения смешанной культуры в пробе AG-Mix. Анализ пробы AG-Mix демонстрирует существенное увеличение видов бактерий по сравнению с первичной культурой в пробе D-MC, что является следствием появления в ранге класса и ниже дополнительных таксонов (рис. 2 и 3).
Причиной перераспределения таксономического состава популяции бактерий из первичных культур после экспонирования с образцами материалов является изменение химического состава исходной минеральной среды в результате перехода органических и минеральных компонентов материалов в растворимое или коллоидное состояние в течение длительного времени. Кроме того, не исключены различного рода взаимодействия бактерий с поверхностными и подповерхностными участками материалов, включая биопленки или другие структуры. Эти обстоятельства приводят к наращиванию ранее некультивируемых и/или плохо растущих в исходной среде бактерий. В то же время происходит подавление роста или элиминация тех штаммов, которые не способны выжить в данных условиях внешней среды при тотальном сокращении общего микробного числа в пробах.
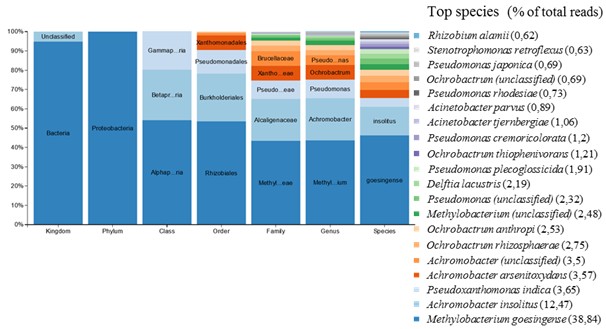
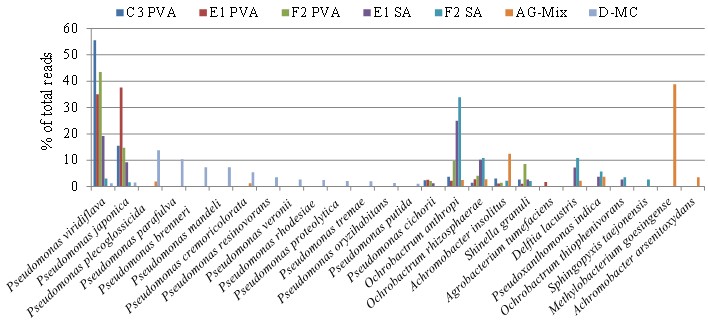
На заключительных этапах отбора отдельные культуры, полученные в результате экспонирования с соответствующими материалами, переносили в базовую минеральную среду с добавлением в качестве единственного источника углерода поливинилового спирта или стирол-акриловой дисперсии. Применение указанных субстратов способствовало дальнейшей дифференциации культур и их обогащению наиболее активными фенотипами (рис. 4; см. приложение, рис. 6). С учетом того факта, что химически и структурно эти субстраты абсолютно разные, они позволяют выявить материал, вносящий наибольший вклад в таксономическое разнообразие культуры, в пробе AG-Mix.
В результате переноса культур после экспонирования с материалами в среду с поливиниловым спиртом происходит обогащение сообщества бактерий изначально мало представленными и плохо культивируемыми штаммами Pseudomonas, в то время как в присутствии стирол-акрилатов увеличивается представленность штаммов Ochrobactrum, Delftia, Pseudoxanthomonas, Achromobacter и снижается изначально высокая концентрация штаммов Pseudomonas. Появление в питательной среде стирол-акрилатов в составе с анионными эмульгаторами модифицирует среду,смещая фокус отбора на другие таксоны бактерий. Это, в частности, объясняет обогащение пробы AG-Mix видами бактерий, представленность которых увеличилась после экспонирования первичных культур из пробы D-MC с материалом Акрэмос-140.
Штамм Methylobacterium goesingense,высоко представленный в пробе AG-Mix (рис. 2 и 3), при наращивании на соответствующих субстратах практически полностью элиминируется (рис. 4; см. приложение, рис. 6). Это, по-видимому, связано с отсутствием метаболических компетенций в отношении использованных субстратов и возросшей конкуренцией со стороны других штаммов, подавляющих его рост.
На рис. 5 представлена сводная диаграмма наиболее представленных бактериальных штаммов в культурах, полученных в результате отбора.
Для получения чистых культур наиболее активных штаммов Ps. viridiflava и/или Ps. japonica выбрана бактериальная культура из пробы C3 PVA, полученная в результате наращивания в поливиниловом спирте бактерий, выделенных после экспозиции первичных культур с отвержденной стирол-акриловой дисперсией Акрэмос-140.
Штамм Ps. japonicaотдельно идентифицирован путем секвенирования полноразмерной последовательности гена 16S рРНК, полученной из чистой культуры. Данные о нуклеотидной последовательности доступны в GeneBank NCBI (Accession number: PQ432336).
Для получения данных о показателях роста на двух видах топлив использовали чистые культуры штаммов Ps. plecoglossicida и Ps. japonica (см. приложение, рис. 7), выделенные до и после экспонирования с материалами соответственно (рис. 6).
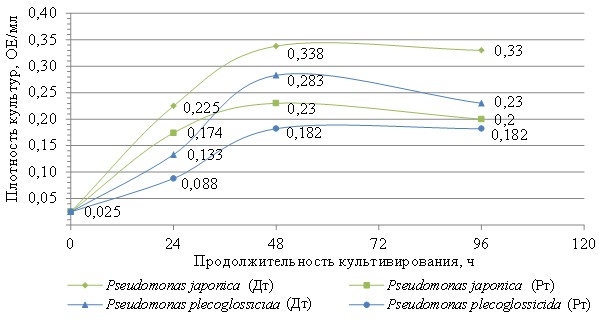
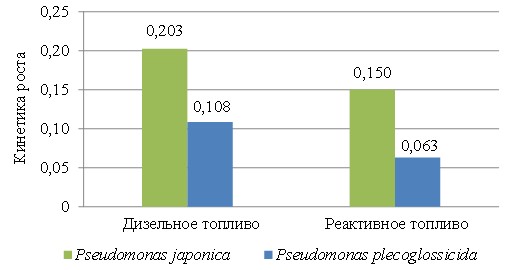
Показано более чем двукратное превышение скорости роста штамма Ps. japonica на реактивном топливе и двукратное – на дизельном топливе по сравнению со штаммом Ps. plecoglossicida (рис. 7).
Полученные данные хорошо согласуются с результатами секвенирования пробы D-MC (рис. 1), в которой штамм Ps. plecoglossicida изначально является одним из доминирующих. На следующих этапах отбора во всех пробах (рис. 1–4; см. приложение, рис. 6) можно наблюдать снижение концентрации этого штамма одновременно с увеличением концентрации других штаммов Pseudomonas, главным образом Ps. viridiflava и Ps. japonica. Это наблюдение свидетельствует о том, что полученные виды бактерий не являются результатом накопления мутаций (мутагенеза) в процессе длительного экспонирования с материалами, поскольку перечисленные штаммы изначально находились в смеси в равных условиях и имели некоторые шансы накопить при экспозиции полезные мутации. При этом у штамма Ps. plecoglossicida, по-видимому, таких шансов было кратно больше, с учетом его концентрации в пробе D-MC (13,76 %), по сравнению с Ps. japonica и Ps. viridiflava (1,56 и 1,21 % соответственно).
Полученные данные и результаты их анализа позволяют также констатировать, что ПЦР-исследование как способ поиска желаемых бактериальных фенотипов в таксономически неоднородных сообществах микроорганизмов весьма трудоемко, обладает низкой эффективностью и более пригодно для детального анализа отдельных видов бактерий.
Заключения
Представленный метод может быть полезен для получения из сложных эндемических сообществ микроорганизмов редких фенотипов бактерий, в том числе биодеструкторов, некультивируемых или плохо культивируемых в богатых питательных средах. Гибкость в выборе субстратов и материалов обеспечивает достаточный уровень масштабируемости – предложенный метод может применяться не только в отношении штаммов Pseudomonas, но и для поиска желаемых фенотипов бактерий в других таксонах. Некоторые из полученных таким способом микроорганизмов могут оказаться естественными продуцентами ценных биологически активных веществ, в том числе биоцидных препаратов, найти применение в методиках испытаний топлив на стойкость к воздействию микроорганизмов, применяться для утилизации материалов или устранения последствий загрязнения нефтепродуктами окружающей среды.
Показатель | Значение показателя для воды водоблока | |||||
1 | 2 | 3 | 4 (I система) | 4 (II система) | АБХМ | |
Содержание, мг/л: нефтепродуктов железа взвешенных веществ хлорид-иона сульфат-иона фосфат-ионов |
0,21 0,161 10 91 131 0,24 |
0,15 0,195 11 112 128 0,05 |
0,09 0,3728 289 277 0,09 |
5,49 0,590 16 187 199 0,05 |
5,40 0,610 19 187 388 0,05 |
0,36 0,351 8 233 430 0,42 |
| Содержание кальция, ммоль/л | 5,5 | 6,6 | 12,4 | 7,9 | 9,1 | 12,2 |
| рН | 8,5 | 8,0 | 8,2 | 8,5 | 8,6 | 8,6 |
| Электропроводность, мкСм/см | 1020 | 1290 | 2580 | 1880 | 1870 | 2250 |
| Жесткость общая, мг-экв/л | 7,2 | 9,6 | 19,8 | 12,3 | 12,6 | 16,4 |
| Щелочность общая, мг-экв/л | 3,9 | 4,2 | 4,2 | 3,5 | 3,4 | 5,4 |
| Общее микробное число, КОЕ/мл | 103 | |||||
| Примечание. В водоблоке 1 (I система) обслуживается установка «Висбрекинг»; в водоблоке 2 (II система) – УПВ-20; в водоблоке 3 (II система) – АКС; в водоблоке 4 (I система) – Л-24-5, Л-24-7, Л-35-5, Л-24-300, «Изориформинг», ЭЛОУ-АВТ-6, «Элементарная сера», Г-43-107М/1, ГОБКК, парк метанола; в водоблоке 4 (II система) – Л-24-5, Л-24-7, Л-35-5, ЦВК-3, Г-43-107М/1, ГОБКК, конденсатная станция; в водоблоке АБХМ (II система) – абсорбционная холодильная машина. | ||||||
Показатель | Значение показателя для воды водоблока | |||
2 | 3 | 4 | 5 | |
Содержание, мг/л: нефтепродуктов железа взвешенных веществ хлорид-иона сульфат-иона фосфат-ионов |
0,76 1,79 17 383 473 1,48 |
0,50 0,66 11 240 298 1,37 |
0,38 0,81 15 419 471 2,33 |
0,60 0,73 7,9 254 338 1,25 |
| Содержание кальция, ммоль/л | 234,46 | 144,29 | 232,46 | 156,3 |
| рН | 8,45 | 8,55 | 8,68 | 8,51 |
| Электропроводность, мкСм/см | 2440 | 1676 | 2005 | 1605 |
| Жесткость общая, мг-экв/л | 15,3 | 10,8 | 16,4 | 10,8 |
| Щелочность общая, мг-экв/л | 6,6 | 4,1 | 6,5 | 4,0 |
| Общее микробное число, КОЕ/мл | 103 | |||
| Примечание. Обслуживаемые установки – потребители охлажденной воды. Водоблок 2 обслуживает: цех 2-3-5/III для производства концентрированного этилена, пропан-пропиленовой фракции, концентрированного пропилена, бутилен-бутадиеновой фракции, жидких продуктов пиролиза; цех 603-609 для производства изопропилбензола; цех 101-615 для производства фенола и ацетона; цех 106-605 для производства инертных газов и сжатого воздуха, холодильного и жидкого аммиака; товарное производство, обеспечивающее прием, хранение, приготовление и откачку сырья, готовой продукции, реагентов и вспомогательных материалов. Водоблок 3 обслуживает: цех 1/I для пиролиза нефтяных углеводородов, компримирования пирогаза, сепарации пирогаза и бутилен-бутадиеновой фракции; цех 1/II для производства олефинов (этилена и пропилена) методом термического расщепления углеводородов нефти; цех 2 для фракционирования пирогаза методом абсорбционной дистилляции, производства концентрированного этилена методом дистилляции и обеспечения соответствующих установок промышленным холодом (пропан-пропиленовая фракция, аммиак). Водоблок 4 обслуживает: цех 202-205 для производства полиэтилена низкой плотности высокого давления в трубчатых реакторах; цех 500-504 для производства полиэтилена низкой плотности высокого давления в автоклавных реакторах; производство полипропилена. Водоблок 5 обслуживает: цех М-1-2 для получения α-метилстирола; цех 400 для производства синтетического этиленпропилендиенового каучука; отделение 8001 для концентрирования пропилена; отделение 405-406 для компримирования газа. | ||||
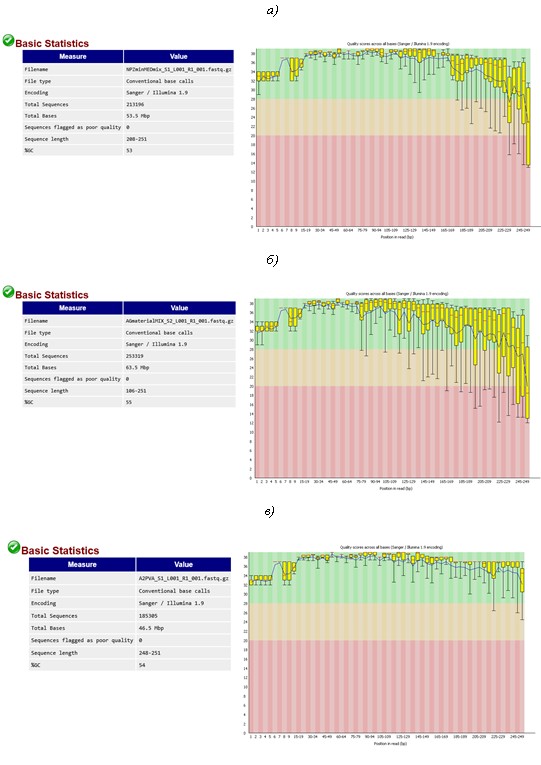
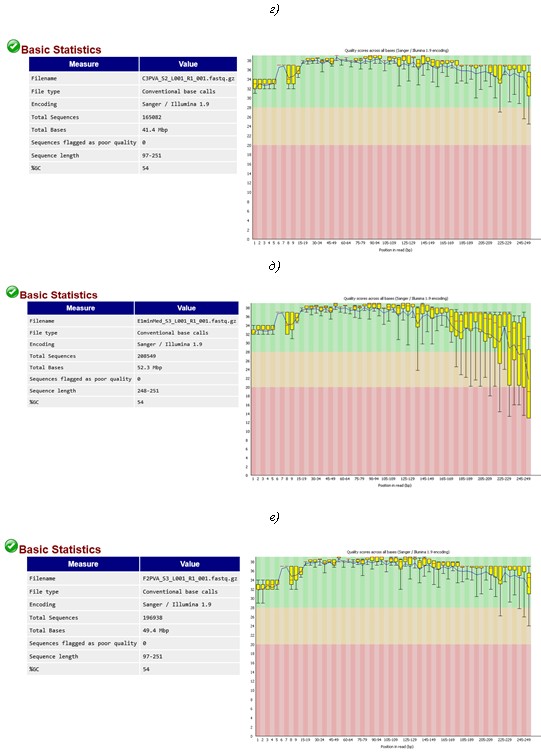
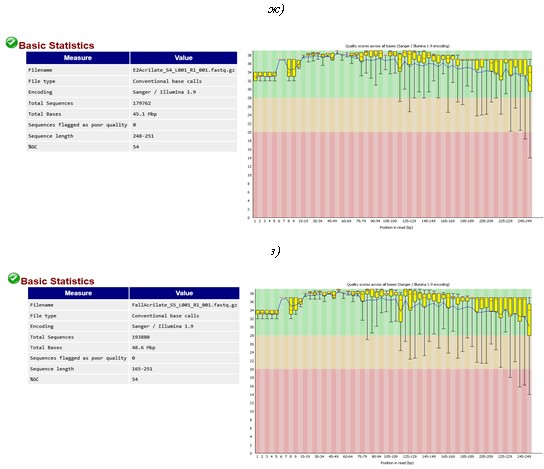
C3 PVA (г), E1 PVA (д), F2 PVA (е), E1 SA (ж) и F2 SA (з). Показатель «Qquality score» (Q) показывает качество прочтений каждого нуклеотида во всех отсеквенированных фрагментах ДНК участка гена 16S рРНК бактерий. Качество прочтений рассчитывали по формуле Q = 10log10P, где P – вероятность ошибки секвенирования. Например, если Р = 0,001, то Q = 30. Это означает, что только 1 нуклеотид из 1000 может быть встроен в цепь ДНК ошибочно. Поэтому Q =30 является пороговым значением для результатов секвенирования с высоким уровнем доверия. Желтые маркеры показывают интервал значений Q по всем прочтениям выборки, красные метки – среднее значение Q, рассчитанное на основе интервала значений Q. Черные «хвосты» указывают на теоретически возможный разброс значений с учетом полученных интервальных значений Q. Перечисленные параметры являются результатом более сложных расчетов, основанных на Q-весовой матрице, откалиброванной производителем, согласно модели секвенатора и передаваемой программе FastQC в соответствующей кодировке, в данном случае – Sanger/Illumina 1.9
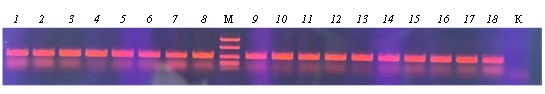
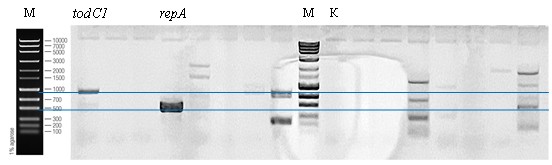
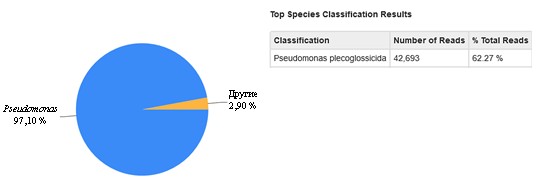

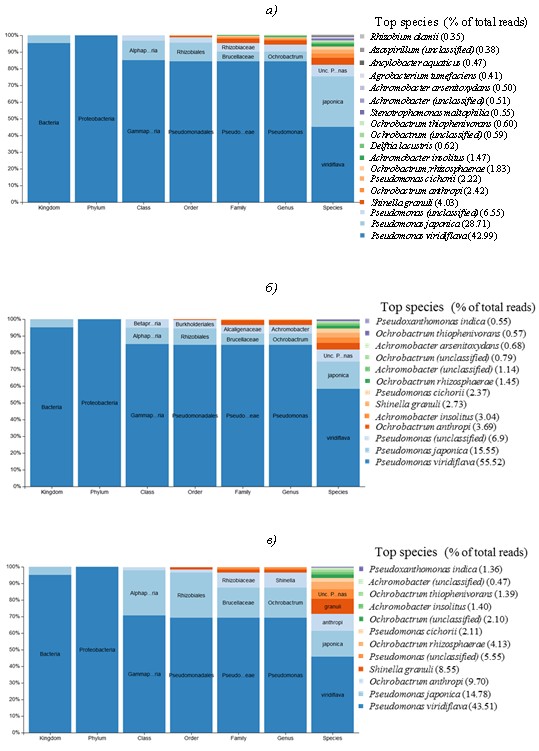
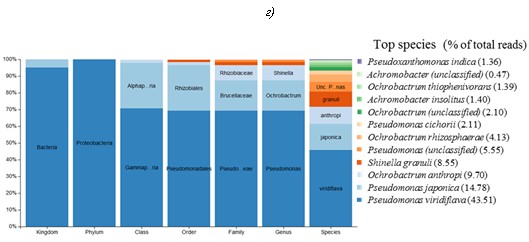
Рис. 6. Видовые составы проб A2 PVA (а), C3 PVA (б), F2 PVA (в) и F2 SA (г) с указанием абсолютного количества прочтений V4-фрагмента 16S рРНК в процессе секвенирования (% of total reads)
- Narancic T., Kevin E. Plastic waste as a global challenge: are biodegradable plastics the answer to the plastic waste problem // Microbiology. 2019. Vol. 165. P. 129–137. DOI: 10.1099/mic.0.000749.
- Sinha V., Patel M.R., Patel J.V. Pet Waste Management by Chemical Recycling: A Review // Journal of Polymers and the Environment. 2010. No. 18. P. 8–25. DOI: 10.1007/s10924-008-0106-7.
- Kablov E.N., Startsev V.O. Climatic aging of aviation polymer composite materials. II. Development of methods for studying the early stages of aging // Russian metallurgy (Metally). 2020. Vol. 2020. No. 10. P. 1088–1094. DOI: 10.1134/S0036029520100110.
- Каблов Е.Н., Старцев В.О. Системный анализ влияния климата на механические свойства полимерных композиционных материалов по данным отечественных и зарубежных источников (обзор) // Авиационные материалы и технологии. 2018. № 2 (51). С. 47–58. DOI: 10.18577/2071-9140-2018-0-2-47-58.
- Каблов Е.Н., Лаптев А.Б., Прокопенко А.Н., Гуляев А.И. Релаксация полимерных композиционных материалов под длительным действием статической нагрузки и климата (обзор). Часть 1. Связующие // Авиационные материалы и технологии. 2021. № 4 (65). Ст. 08. URL: http://www.journal.viam.ru (дата обращения: 29.03.2024). DOI: 10.18577/2713-0193-2021-0-4-70-80.
- Каблов Е.Н. Материалы нового поколения и цифровые технологии их переработки // Вестник Российской академии наук. 2020. Т. 90. № 4. С. 331–334.
- Startsev O.V., Krotov A., Mashinskaya G. Climatic Ageing of Organic Fiber Reinforced Plastics: Water Effect // Journal Polymeric Material. 1997. Vol. 37. Р. 161–171.
- Frisch H.L. Diffusion in polymers // Journal of Applied Polymer Science. 1970. No. 14. P. 1657.
- Lucas N., Bienaime C., Belloy C. et al. Polymer biodegradation: mechanisms and estimation techniques // Chemosphere. 2008. No. 4. P. 429. DOI: 10.1016/j.chemosphere.2008.06.064.6.
- Eskander S., Saleh H. Biodegradation: Process Mechanism // Environmental Science and Engineering. 2017. Vol. 8. No. 1. P. 1–31.
- Ермишев В.Ю., Лаптев А.Б., Старцев В.О. Особенности оценки стойкости полимерных материалов к биодеструкции в лабораторных условиях. Часть 1. Разрушение полимерных материалов в природных средах, выбор штаммов бактерий, питательные среды и условия культивирования // Труды ВИАМ. 2023. № 7 (125). Ст. 12. URL: http://www.viam-works.ru (дата обращения: 29.03.2024). DOI: 10.18577/2307-6046-2023-0-7-138-148.
- Лаптев А.Б., Павлов М.Р., Новиков А.А., Славин А.В. Современные тенденции развития испытаний материалов на стойкость к климатическим факторам (обзор). Часть 1. Испытания новых материалов // Труды ВИАМ. 2021. № 1 (95). Ст. 12. URL: http://www.viam-works.ru (дата обращения: 29.03.2024). DOI: 10.18577/2307-6046-2021-0-1-114-122.
- Yoshida S., Hiraga K., Takehana T. et al. A bacterium that degrades and assimilates poly(ethyleneterephthalate) // Science. 2016. Vol. 353. P. 759–769.
- Ермишев В.Ю. Метаболические возможности бактерий в отношении синтетических углеводородов, используемых в производстве неметаллических материалов (обзор) // Труды ВИАМ. 2023. № 2 (120). Ст. 11. URL: http://www.viam-works.ru (дата обращения: 29.03.2024). DOI: 10.18577/2307-6046-2023-0-2-132-146.
- Baker G.C., Smith J.J., Cowan D.A. Review and re-analysis of domain-specific 16S primers // Journal of Microbiological Methods. 2003. Vol. 55. P. 541–555. DOI: 10.1016/j.mimet.2003.08.009.
- Polz M.F., Cavanaugh C.M. Bias in template-to-product ratios in multitemplate PCR // Applied and Environmental Microbiology. 1998. Vol. 11. P. 3724–3730. DOI: 10.1128/AEM.64.10.3724-3730.1998.
- Okonechnikov K., Golosova O., Fursov M. A unified bioinformatics toolkit // Bioinformatics. 2012. Vol. 28. P. 1166–1167. DOI: 10.1093/bioinformatics/bts091.
- Vashishtha K., Gaud C., Andrews S., Krueger C. A quality control tool to analyse sequencing library compositions // F1000Research. 2022. Vol. 11. P. 1122. DOI: 10.12688/f1000research.125325.2
- Nair A.V., Pradeep M.A., Vijayan K.K. Molecular approach for the rapid detection of Bacillus and Pseudomonas genera dominant antagonistic groups from diverse ecological niches using colony multiplex PCR // Journal of Industrial Microbiology and Biotechnology. 2014. Vol. 41. P. 1367–5435. DOI: 10.1007/s10295-014-1441-4.
- Rainey P. A pair of PCR primers for Incp-9 plasmids // Microbiology. 1999. Vol. 145. P. 11. DOI: 10.1099/00221287-145-11-3003.
- Baldwin B., Nakatsu C., Nies L. Detection and Enumeration of Aromatic Oxygenase Genes by Multiplex and Real-Time PCR // Applied and Environmental Microbiology. 2003. Vol. 69. P. 3350–3358. DOI: 10.1128/AEM.69.6.3350-3358.2003.